

Activation and modulation of concatemeric GABA-A receptors expressed in human embryonic kidney cells
- PMID: 19289569
- PMCID: PMC2684887
- DOI: 10.1124/mol.108.054510
Activation and modulation of concatemeric GABA-A receptors expressed in human embryonic kidney cells
Abstract
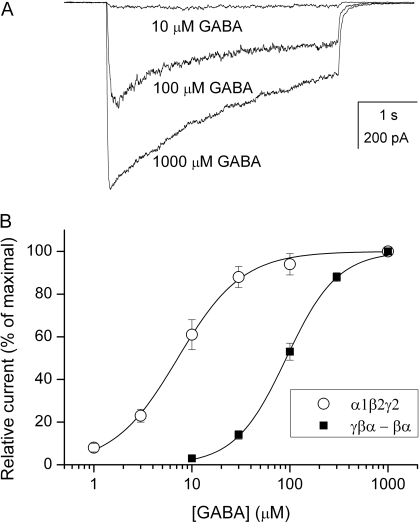
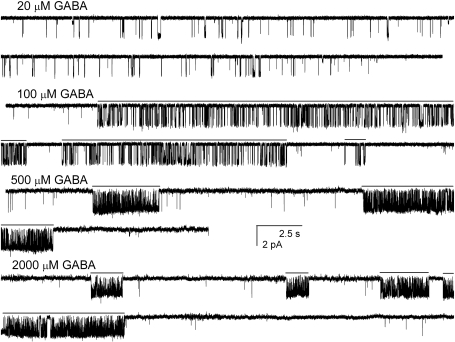
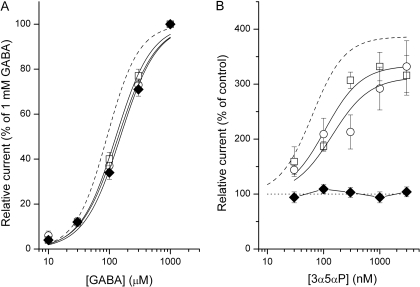
We have employed whole-cell and single-channel electrophysiology to examine the kinetic and pharmacological properties of GABA-A receptors consisting of gamma2L-beta2-alpha1 and beta2-alpha1 subunit concatemeric constructs expressed in human embryonic kidney cells. Concatemeric receptors activated by GABA exhibited the same single-channel conductance, channel opening rate constant, and basic open- and closed-time properties as receptors containing free subunits. However, the whole-cell GABA dose-response and the single-channel effective opening rate curves were shifted to higher GABA concentrations, suggesting that the concatemeric receptors have a lower affinity to GABA. Pharmacological tests demonstrated that the concatemeric receptors were potentiated by pentobarbital, diazepam, and the neurosteroid (3alpha,5alpha)-3-hydroxypregnan-20-one (3alpha5alphaP), and were insensitive to Zn(2+). Selective introduction of the alpha1Q241L mutation, previously shown to abolish alpha1beta2gamma2L channel potentiation by neurosteroids, into one of the two concatemeric constructs had a relatively small effect on receptor activation by GABA or macroscopic potentiation by the neurosteroid 3alpha5alphaP. Single-channel measurements showed that the kinetic mechanism of action of the steroid is unchanged when the mutation is introduced to the gamma2L-beta2-alpha1 concatemer. We infer that a single wild-type alpha subunit is capable of mediating the full set of kinetic effects in the presence of steroids. Introduction of the alpha1Q241W mutation, previously shown to mimic the effect of the steroid on alpha1beta2gamma2L channels, selectively into either concatemeric construct altered the mode of activity elicited by P4S, but the presence of mutations in both alpha subunits was required to affect open-time distributions. The data indicate that the alpha1Q241W mutation acts as a partial steroid modulator.
Figures





References
-
- Baumann SW, Baur R, and Sigel E (2001) Subunit arrangement of γ-aminobutyric acid type A receptors. J Biol Chem 276 36275-36280. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
