

Elevated P75NTR expression causes death of engrailed-deficient midbrain dopaminergic neurons by Erk1/2 suppression
- PMID: 19291307
- PMCID: PMC2667502
- DOI: 10.1186/1749-8104-4-11
Elevated P75NTR expression causes death of engrailed-deficient midbrain dopaminergic neurons by Erk1/2 suppression
Abstract
Background: The homeodomain transcription factors Engrailed-1 and Engrailed-2 are required for the survival of mesencephalic dopaminergic (mesDA) neurons in a cell-autonomous and gene-dose-dependent manner. Homozygote mutant mice, deficient of both genes (En1-/-;En2-/-), die at birth and exhibit a loss of all mesDA neurons by mid-gestation. In heterozygote animals (En1+/-;En2-/-), which are viable and fertile, postnatal maintenance of the nigrostriatal dopaminergic system is afflicted, leading to a progressive degeneration specific to this subpopulation and Parkinson's disease-like molecular and behavioral deficits.
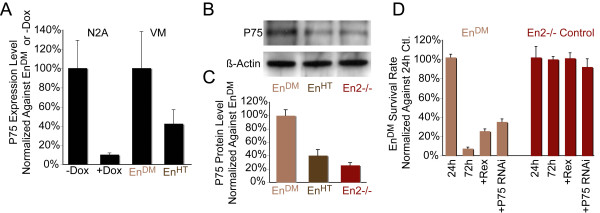
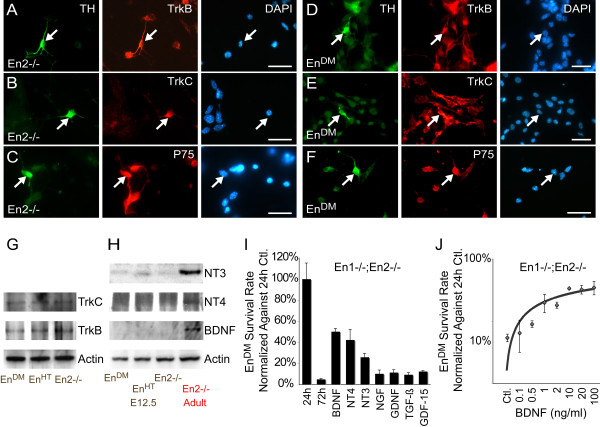
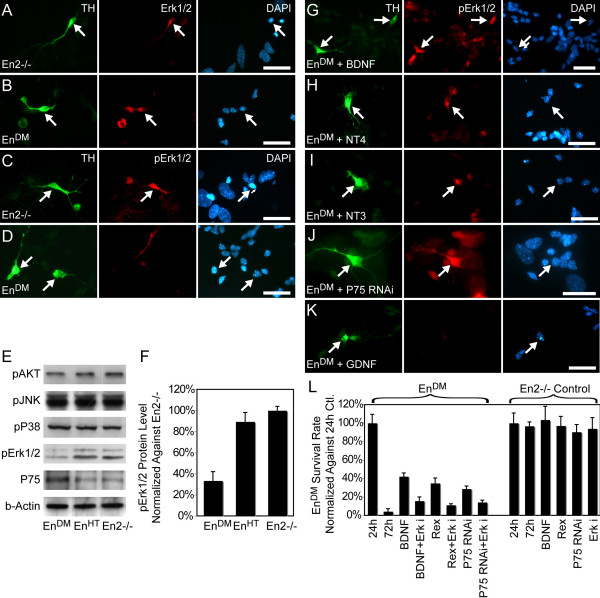
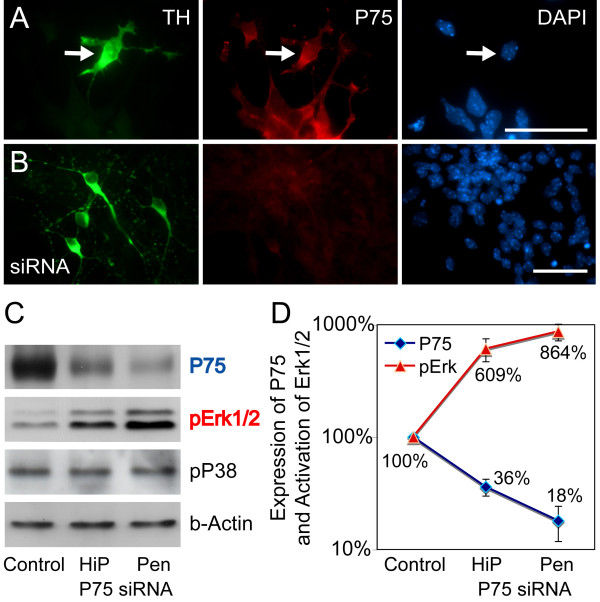
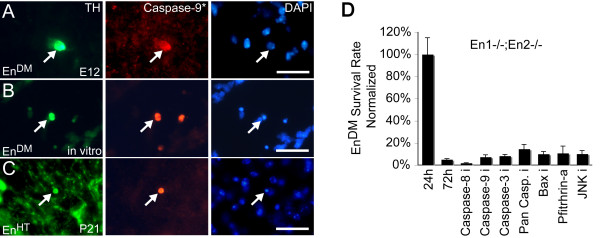
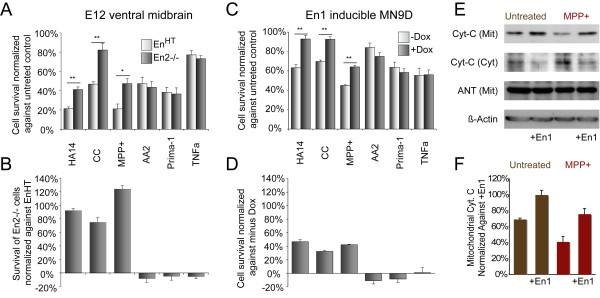
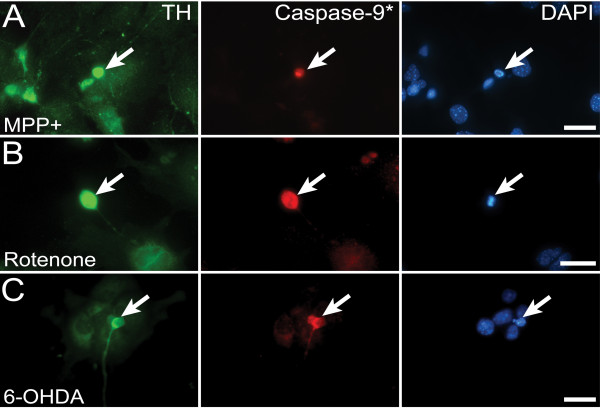
Results: In this work, we show that the dose of Engrailed is inversely correlated to the expression level of the pan-neurotrophin receptor gene P75NTR (Ngfr). Loss of mesDA neurons in the Engrailed-null mutant embryos is caused by elevated expression of this neurotrophin receptor: Unusually, in this case, the cell death signal of P75NTR is mediated by suppression of Erk1/2 (extracellular-signal-regulated kinase 1/2) activity. The reduction in expression of Engrailed, possibly related to the higher levels of P75NTR, also decreases mitochondrial stability. In particular, the dose of Engrailed determines the sensitivity to cell death induced by the classic Parkinson-model toxin MPTP and to inhibition of the anti-apoptotic members of the Bcl-2 family of proteins.
Conclusion: Our study links the survival function of the Engrailed genes in developing mesDA neurons to the regulation of P75NTR and the sensitivity of these neurons to mitochondrial insult. The similarities to the disease etiology in combination with the nigral phenotype of En1+/-;En2-/- mice suggests that haplotype variations in the Engrailed genes and/or P75NTR that alter their expression levels could, in part, determine susceptibility to Parkinson's disease.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
