

Regulation of the processivity and intracellular localization of Saccharomyces cerevisiae dynein by dynactin
- PMID: 19293377
- PMCID: PMC2657088
- DOI: 10.1073/pnas.0900976106
Regulation of the processivity and intracellular localization of Saccharomyces cerevisiae dynein by dynactin
Abstract
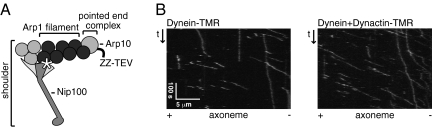
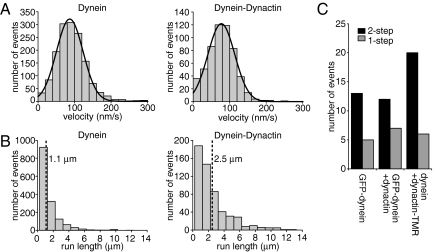
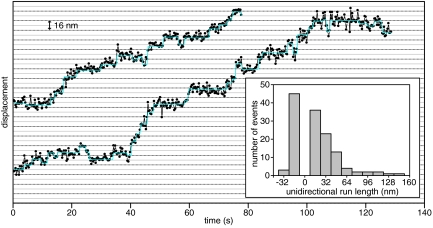
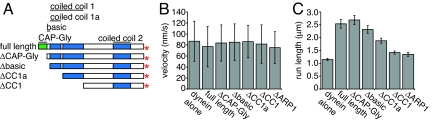
Dynactin, a large multisubunit complex, is required for intracellular transport by dynein; however, its cellular functions and mechanism of action are not clear. Prior studies suggested that dynactin increases dynein processivity by tethering the motor to the microtubule through its own microtubule binding domains. However, this hypothesis could not be tested without a recombinant source of dynactin. Here, we have produced recombinant dynactin and dynein in Saccharomyces cerevisiae, and examined the effect of dynactin on dynein in single-molecule motility assays. We show that dynactin increases the run length of single dynein motors, but does not alter the directionality of dynein movement. Enhancement of dynein processivity by dynactin does not require the microtubule (MT) binding domains of Nip100 (the yeast p150(Glued) homolog). Dynactin lacking these MT binding domains also supports the proper localization and function of dynein during nuclear segregation in vivo. Instead, a segment of the coiled-coil of Nip100 is required for these activities. Our results directly demonstrate that dynactin increases the processivity of dynein through a mechanism independent of microtubule tethering.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Karki S, Holzbaur EL. Cytoplasmic dynein and dynactin in cell division and intracellular transport. Curr Opin Cell Biol. 1990;11:45–53. - PubMed
-
- Schroer TA. Dynactin. Annu Rev Cell Dev Biol. 2004;20:759–779. - PubMed
-
- Puls I, et al. Mutant dynactin in motor neuron disease. Nat Genet. 2003;33:455–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
