

Lnk inhibits myeloproliferative disorder-associated JAK2 mutant, JAK2V617F
- PMID: 19293402
- PMCID: PMC2698587
- DOI: 10.1189/jlb.0908575
Lnk inhibits myeloproliferative disorder-associated JAK2 mutant, JAK2V617F
Abstract
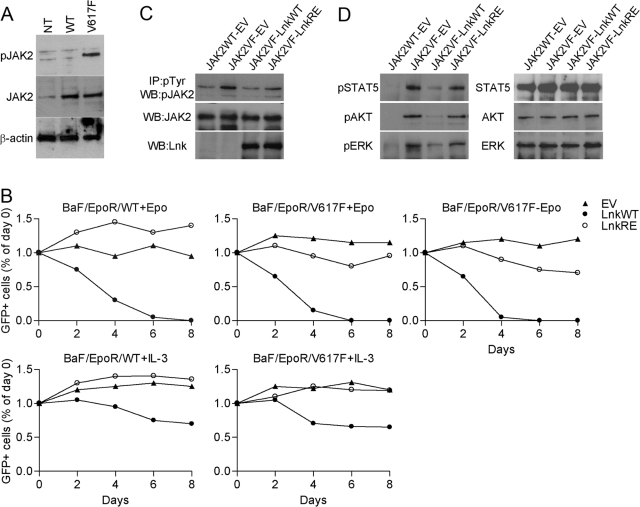
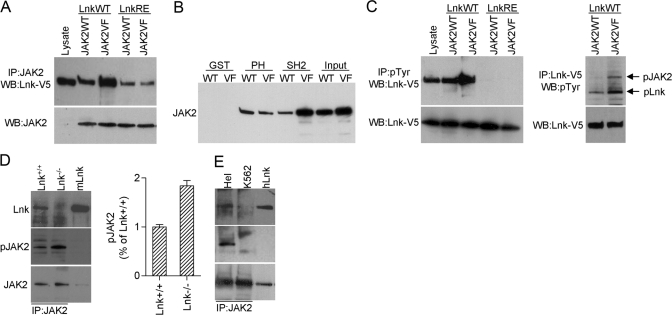
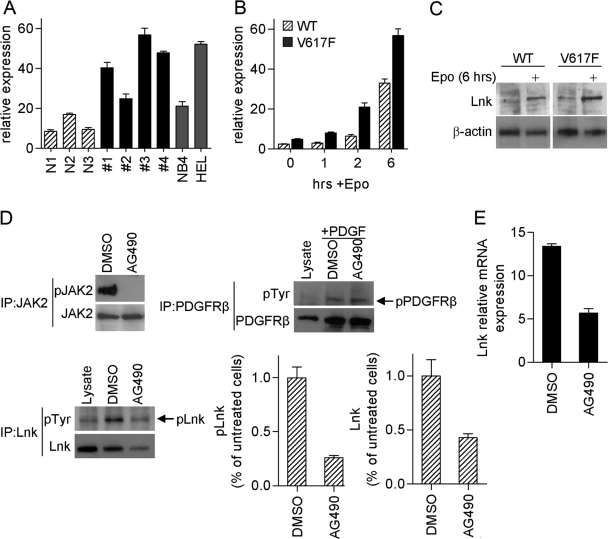
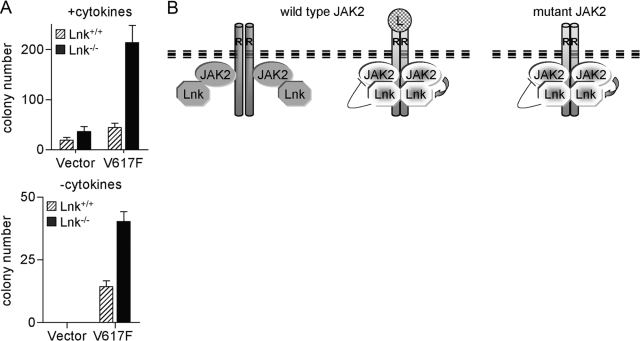
The JAK2 mutation JAK2V617F is found frequently in patients with myeloproliferative disorders (MPD) and transforms hematopoietic cells to cytokine-independent proliferation when expressed with specific cytokine receptors. The Src homology 2 (SH2) and pleckstrin homology (PH) domain-containing adaptor protein Lnk (SH2B3) is a negative regulator of hematopoietic cytokine signaling. Here, we show that Lnk is a potent inhibitor of JAK2V617F constitutive activity. Lnk down-regulates JAK2V617F-mediated signaling and transformation in hematopoietic Ba/F3-erythropoietin receptor cells. Furthermore, in CFU assays, Lnk-deficient murine bone marrow cells are significantly more sensitive to transformation by JAK2V617F than wild-type (WT) cells. Lnk, through its SH2 and PH domains, interacts with WT and mutant JAK2 and is phosphorylated by constitutively activated JAK2V617F. Finally, we found that Lnk levels are high in CD34(+) hematopoietic progenitors from MPD patients and that Lnk expression is induced following JAK2 activation. Our data suggest that JAK2V617F is susceptible to endogenous negative-feedback regulation, providing new insights into the molecular pathogenesis of MPD.
Figures
References
-
- Parganas E, Wang D, Stravopodis D, Topham D J, Marine J C, Teglund S, Vanin E F, Bodner S, Colamonici O R, van Deursen J M, Grosveld G, Ihle J N. Jak2 is essential for signaling through a variety of cytokine receptors. Cell. 1998;93:385–395. - PubMed
-
- Campbell P J, Green A R. The myeloproliferative disorders. N Engl J Med. 2006;355:2452–2466. - PubMed
-
- Levine R L, Pardanani A, Tefferi A, Gilliland D G. Role of JAK2 in the pathogenesis and therapy of myeloproliferative disorders. Nat Rev Cancer. 2007;7:673–683. - PubMed
-
- Baxter E J, Scott L M, Campbell P J, East C, Fourouclas N, Swanton S, Vassiliou G S, Bench A J, Boyd E M, Curtin N, Scott M A, Erber W N, Green A R. Cancer Genome Project Acquired mutation of the tyrosine kinase JAK2 in human myeloproliferative disorders. Lancet. 2005;365:1054–1061. - PubMed
-
- James C, Ugo V, Le Couedic J P, Staerk J, Delhommeau F, Lacout C, Garcon L, Raslova H, Berger R, Bennaceur-Griscelli A, Villeval J L, Constantinescu S N, Casadevall N, Vainchenker W. A unique clonal JAK2 mutation leading to constitutive signaling causes polycythemia vera. Nature. 2005;434:1144–1148. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
