

Inhibition of p75 neurotrophin receptor attenuates isoflurane-mediated neuronal apoptosis in the neonatal central nervous system
- PMID: 19293698
- PMCID: PMC2767332
- DOI: 10.1097/ALN.0b013e31819b602b
Inhibition of p75 neurotrophin receptor attenuates isoflurane-mediated neuronal apoptosis in the neonatal central nervous system
Abstract
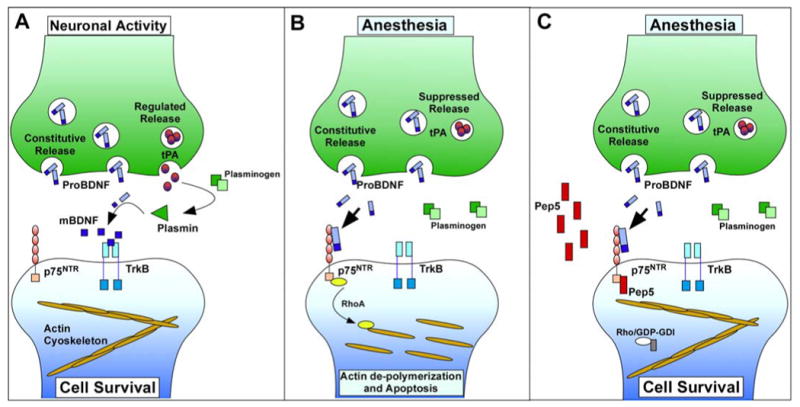
Background: Exposure to anesthetics during synaptogenesis results in apoptosis and subsequent cognitive dysfunction in adulthood. Probrain-derived neurotrophic factor (proBDNF) is involved in synaptogenesis and can induce neuronal apoptosis via p75 neurotrophic receptors (p75). proBDNF is cleaved into mature BDNF (mBDNF) by plasmin, a protease converted from plasminogen by tissue plasminogen activator (tPA) that is released with neuronal activity; mBDNF supports survival and stabilizes synapses through tropomyosin receptor kinase B. The authors hypothesized that anesthetics suppress tPA release from neurons, enhance p75 signaling, and reduce synapses, resulting in apoptosis.
Methods: Primary neurons (DIV5) and postnatal day 5-7 (PND5-7) mice were exposed to isoflurane (1.4%, 4 h) in 5% CO2, 95% air. Apoptosis was assessed by cleaved caspase-3 (Cl-Csp3) immunoblot and immunofluorescence microscopy. Dendritic spine changes were evaluated with the neuronal spine marker, drebrin. Changes in synapses in PND5-7 mouse hippocampi were assessed by electron microscopy. Primary neurons were exposed to tPA, plasmin, or pharmacologic inhibitors of p75 (Fc-p75 or TAT-Pep5) 15 min before isoflurane. TAT-Pep5 was administered by intraperitoneal injection to PND5-7 mice 15 min before isoflurane.
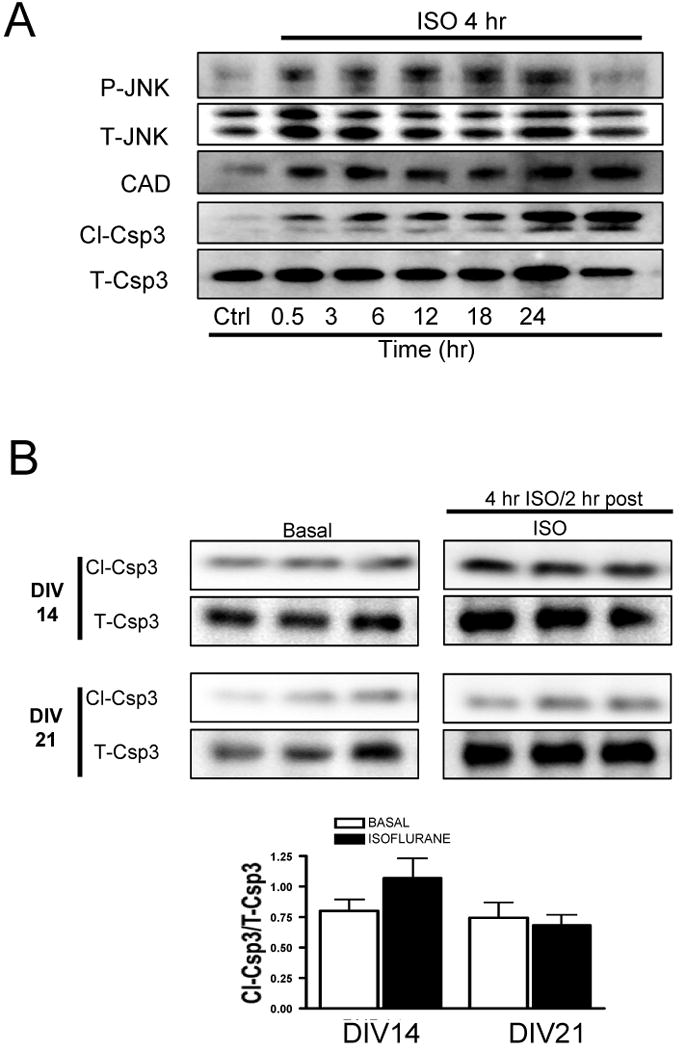
Results: Exposure of neurons in vitro (DIV5) to isoflurane decreased tPA in the culture medium, reduced drebrin expression (marker of dendritic filopodial spines), and enhanced Cl-Csp3. tPA, plasmin, or TAT-Pep5 stabilized dendritic filopodial spines and decreased Cl-Csp3 in neurons. TAT-Pep5 blocked isoflurane-mediated increase in Cl-Csp3 and reduced synapses in PND5-7 mouse hippocampi.
Conclusion: tPA, plasmin, or p75 inhibition blocked isoflurane-mediated reduction in dendritic filopodial spines and neuronal apoptosis in vitro. Isoflurane reduced synapses and enhanced Cl-Csp3 in the hippocampus of PND5-7 mice, the latter effect being mitigated by p75 inhibition in vivo. These data support the hypothesis that isoflurane neurotoxicity in the developing rodent brain is mediated by reduced synaptic tPA release and enhanced proBDNF/p75-mediated apoptosis.
Figures




Comment in
-
Between Clotho and Lachesis: how isoflurane seals neuronal fate.Anesthesiology. 2009 Apr;110(4):709-11. doi: 10.1097/ALN.0b013e31819b590a. Anesthesiology. 2009. PMID: 19276964 No abstract available.
-
Role of p75 neurotrophin receptor in isoflurane-mediated neuronal changes.Anesthesiology. 2009 Nov;111(5):1162-3; author reply 1163-4. doi: 10.1097/ALN.0b013e3181bbc177. Anesthesiology. 2009. PMID: 19858884 No abstract available.
References
-
- Jevtovic-Todorovic V, Wozniak DF, Benshoff ND, Olney JW. A comparative evaluation of the neurotoxic properties of ketamine and nitrous oxide. Brain Res. 2001;895:264–7. - PubMed
-
- Fredriksson A, Ponten E, Gordh T, Eriksson P. Neonatal Exposure to a Combination of N-Methyl-d-aspartate and gamma-Aminobutyric Acid Type A Receptor Anesthetic Agents Potentiates Apoptotic Neurodegeneration and Persistent Behavioral Deficits. Anesthesiology. 2007;107:427–36. - PubMed
-
- Olney JW, Ishimaru MJ, Bittigau P, Ikonomidou C. Ethanol-induced apoptotic neurodegeneration in the developing brain. Apoptosis. 2000;5:515–21. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
