

Distinct role of PLCbeta3 in VEGF-mediated directional migration and vascular sprouting
- PMID: 19295129
- PMCID: PMC2720932
- DOI: 10.1242/jcs.041913
Distinct role of PLCbeta3 in VEGF-mediated directional migration and vascular sprouting
Abstract
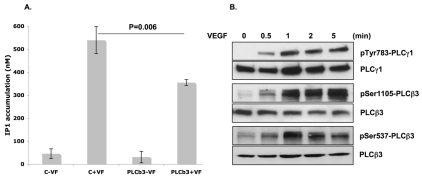
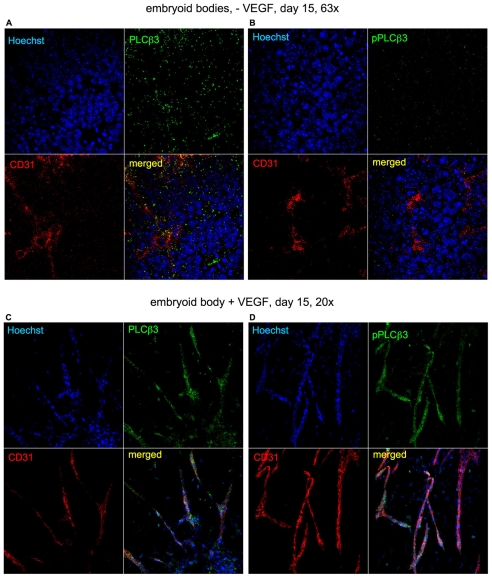
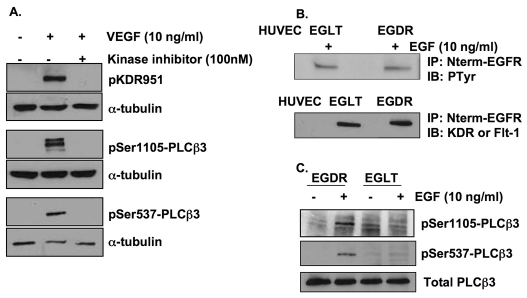
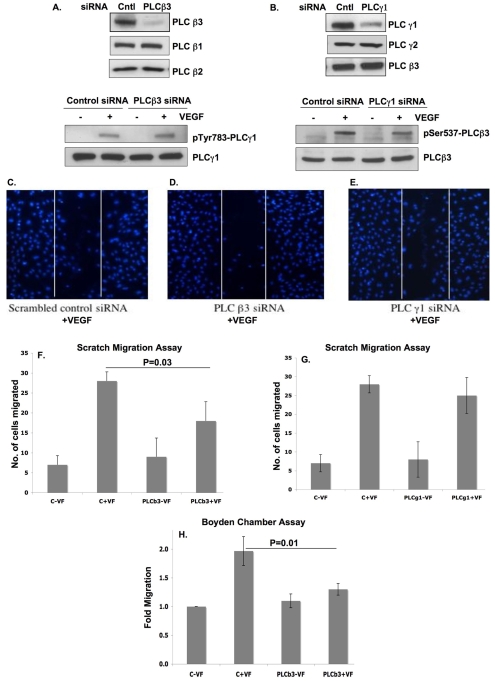
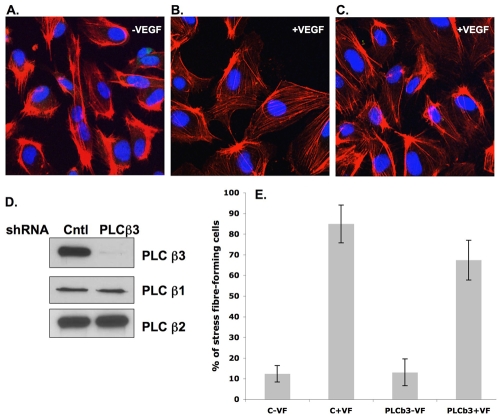
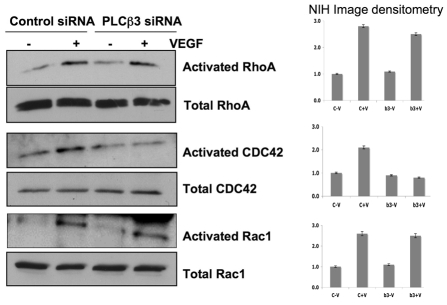
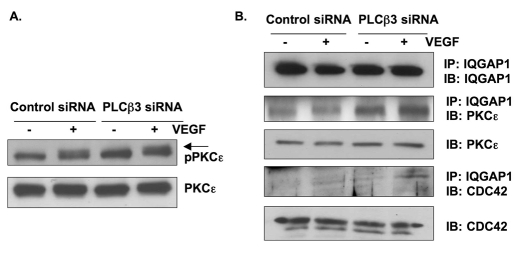
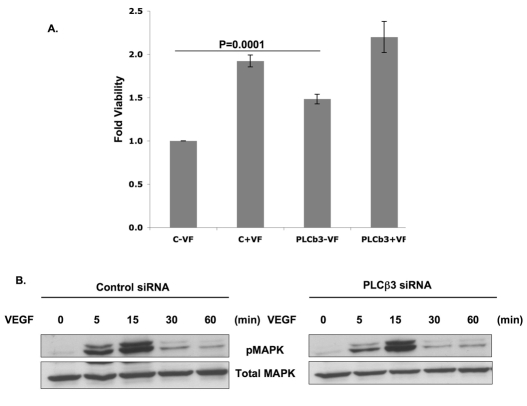
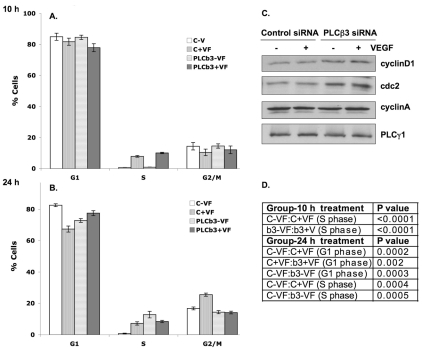
Endothelial cell proliferation and migration is essential to angiogenesis. Typically, proliferation and chemotaxis of endothelial cells is driven by growth factors such as vascular endothelial growth factor (VEGF) and basic fibroblast growth factor (bFGF). VEGF activates phospholipases (PLCs) - specifically PLCgamma1 - that are important for tubulogenesis, differentiation and DNA synthesis. However, we show here that VEGF, specifically through VEGFR2, induces phosphorylation of two serine residues on PLCbeta3, and this was confirmed in an ex vivo embryoid body model. Knockdown of PLCbeta3 in HUVEC cells affects IP3 production, actin reorganization, migration and proliferation; whereas migration is inhibited, proliferation is enhanced. Our data suggest that enhanced proliferation is precipitated by an accelerated cell cycle, and decreased migration by an inability to activate CDC42. Given that PLCbeta3 is typically known as an effector of heterotrimeric G-proteins, our data demonstrate a unique crosstalk between the G-protein and receptor tyrosine kinase (RTK) axes and reveal a novel molecular mechanism of VEGF signaling and, thus, angiogenesis.
Figures
References
-
- Berridge, M. J. (1993). Inositol trisphosphate and calcium signalling. Nature 361, 315-325. - PubMed
-
- Cascone, I., Audero, E., Giraudo, E., Napione, L., Maniero, F., Philips, M. R., Collard, J. G., Serini, G. and Bussolino, F. (2003). Tie-2-dependent activation of RhoA and Rac1 participates in endothelial cell motility triggered by angiopoietin-1. Blood 102, 2482-2490. - PubMed
-
- Dvorak, H. F., Dvorak, A. M., Manseau, E. J., Wiberg, L. and Churchill, W. H. (1979). Fibrin gel investment associated with line 1 and line 10 solid tumor growth, angiogenesis, and fibroplasia in guinea pigs: role of cellular immunity, myofibroblasts, microvascular damage, and infarction in line 1 tumor regression. J. Natl. Cancer Inst. 62, 1459-1472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
