

Cargo and dynamin regulate clathrin-coated pit maturation
- PMID: 19296720
- PMCID: PMC2656549
- DOI: 10.1371/journal.pbio.1000057
Cargo and dynamin regulate clathrin-coated pit maturation
Abstract
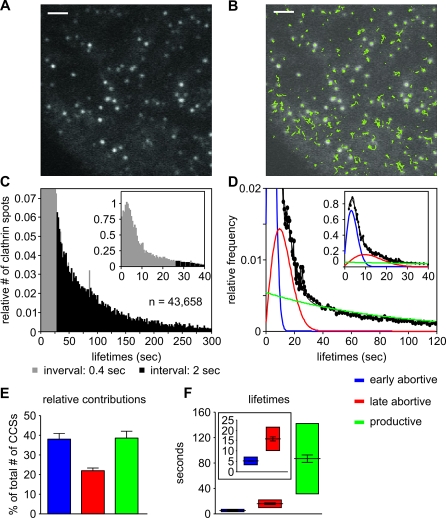
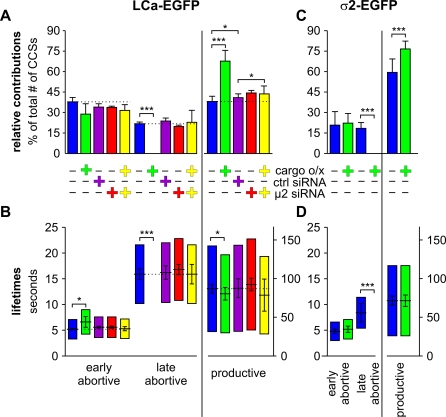
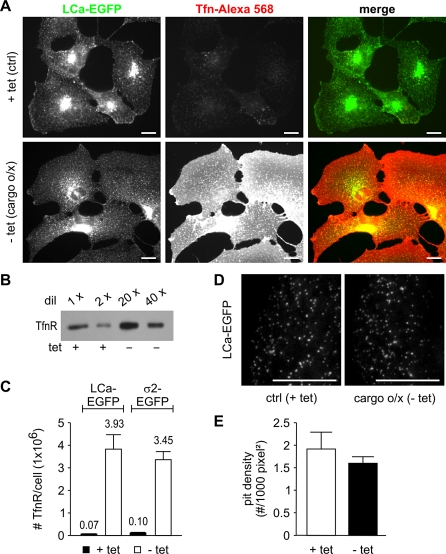
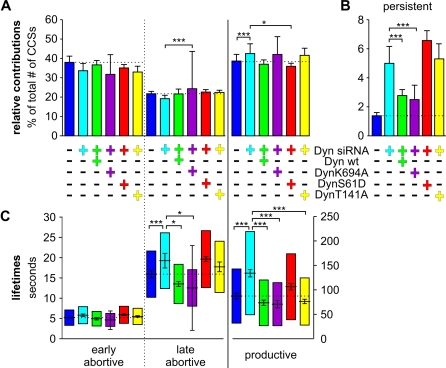
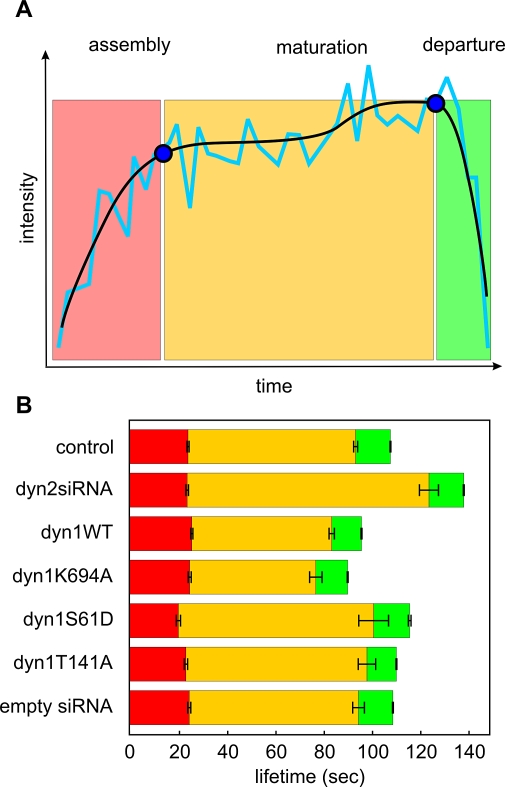
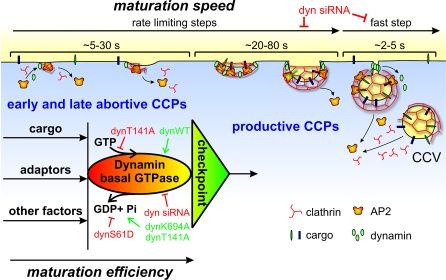
Total internal reflection fluorescence microscopy (TIR-FM) has become a powerful tool for studying clathrin-mediated endocytosis. However, due to difficulties in tracking and quantifying their heterogeneous dynamic behavior, detailed analyses have been restricted to a limited number of selected clathrin-coated pits (CCPs). To identify intermediates in the formation of clathrin-coated vesicles and factors that regulate progression through these stages, we used particle-tracking software and statistical methods to establish an unbiased and complete inventory of all visible CCP trajectories. We identified three dynamically distinct CCP subpopulations: two short-lived subpopulations corresponding to aborted intermediates, and one longer-lived productive subpopulation. In a manner dependent on AP2 adaptor complexes, increasing cargo concentration significantly enhances the maturation efficiency of productive CCPs, but has only minor effects on their lifetimes. In contrast, small interfering RNA (siRNA) depletion of dynamin-2 GTPase and reintroduction of wild-type or mutant dynamin-1 revealed dynamin's role in controlling the turnover of abortive intermediates and the rate of CCP maturation. From these data, we infer the existence of an endocytic restriction or checkpoint, responsive to cargo and regulated by dynamin.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
Similar articles
-
Divergent modes for cargo-mediated control of clathrin-coated pit dynamics.Mol Biol Cell. 2013 Jun;24(11):1725-34, S1-12. doi: 10.1091/mbc.E12-07-0550. Epub 2013 Mar 27. Mol Biol Cell. 2013. PMID: 23536704 Free PMC article.
-
Endocytic accessory proteins are functionally distinguished by their differential effects on the maturation of clathrin-coated pits.Mol Biol Cell. 2009 Jul;20(14):3251-60. doi: 10.1091/mbc.e09-03-0256. Epub 2009 May 20. Mol Biol Cell. 2009. PMID: 19458185 Free PMC article.
-
Cargo regulates clathrin-coated pit dynamics.Cell. 2006 Oct 6;127(1):113-24. doi: 10.1016/j.cell.2006.08.035. Cell. 2006. PMID: 17018281
-
Understanding living clathrin-coated pits.Traffic. 2004 May;5(5):327-37. doi: 10.1111/j.1398-9219.2004.00187.x. Traffic. 2004. PMID: 15086782 Review.
-
Dissecting dynamin's role in clathrin-mediated endocytosis.Biochem Soc Trans. 2009 Oct;37(Pt 5):1022-6. doi: 10.1042/BST0371022. Biochem Soc Trans. 2009. PMID: 19754444 Free PMC article. Review.
Cited by
-
An internally eGFP-tagged α-adaptin is a fully functional and improved fiduciary marker for clathrin-coated pit dynamics.Traffic. 2020 Sep;21(9):603-616. doi: 10.1111/tra.12755. Epub 2020 Jul 22. Traffic. 2020. PMID: 32657003 Free PMC article.
-
u-track3D: Measuring, navigating, and validating dense particle trajectories in three dimensions.Cell Rep Methods. 2023 Dec 18;3(12):100655. doi: 10.1016/j.crmeth.2023.100655. Epub 2023 Dec 1. Cell Rep Methods. 2023. PMID: 38042149 Free PMC article.
-
CALM regulates clathrin-coated vesicle size and maturation by directly sensing and driving membrane curvature.Dev Cell. 2015 Apr 20;33(2):163-75. doi: 10.1016/j.devcel.2015.03.002. Dev Cell. 2015. PMID: 25898166 Free PMC article.
-
Structural basis of an endocytic checkpoint that primes the AP2 clathrin adaptor for cargo internalization.Nat Struct Mol Biol. 2022 Apr;29(4):339-347. doi: 10.1038/s41594-022-00749-z. Epub 2022 Mar 28. Nat Struct Mol Biol. 2022. PMID: 35347313 Free PMC article.
-
Receptor Heterodimerization Modulates Endocytosis through Collaborative and Competitive Mechanisms.Biophys J. 2019 Aug 20;117(4):646-658. doi: 10.1016/j.bpj.2019.07.012. Epub 2019 Jul 16. Biophys J. 2019. PMID: 31358286 Free PMC article.
References
-
- Rappoport JZ, Taha BW, Lemeer S, Benmerah A, Simon SM. The AP-2 complex is excluded from the dynamic population of plasma membrane-associated clathrin. J Biol Chem. 2003;278:47357–47360. - PubMed
-
- Ehrlich M, Boll W, Van Oijen A, Hariharan R, Chandran K, et al. Endocytosis by random initiation and stabilization of clathrin-coated pits. Cell. 2004;118:591–605. - PubMed
-
- Keyel PA, Watkins SC, Traub LM. Endocytic adaptor molecules reveal an endosomal population of clathrin by total internal reflection fluorescence microscopy. J Biol Chem. 2004;279:13190–13204. - PubMed
-
- Merrifield CJ, Perrais D, Zenisek D. Coupling between clathrin-coated-pit invagination, cortactin recruitment, and membrane scission observed in live cells. Cell. 2005;121:593–606. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
