

RacGAP50C directs perinuclear gamma-tubulin localization to organize the uniform microtubule array required for Drosophila myotube extension
- PMID: 19297411
- PMCID: PMC2674253
- DOI: 10.1242/dev.031823
RacGAP50C directs perinuclear gamma-tubulin localization to organize the uniform microtubule array required for Drosophila myotube extension
Abstract
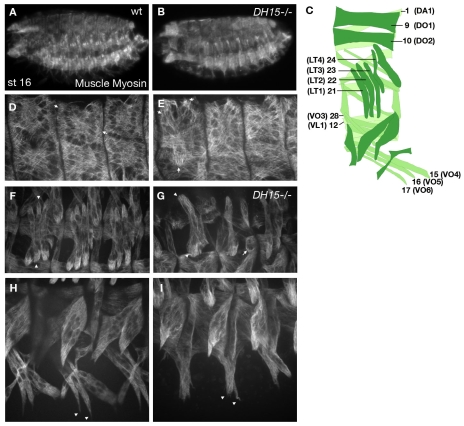
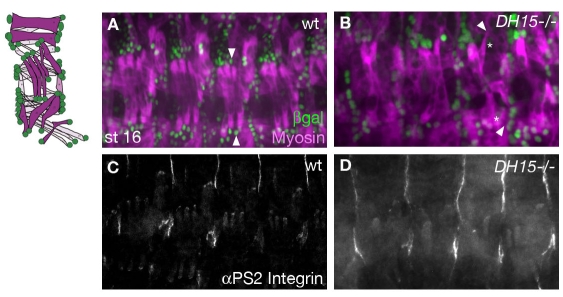
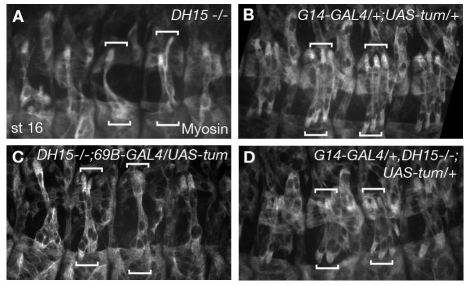
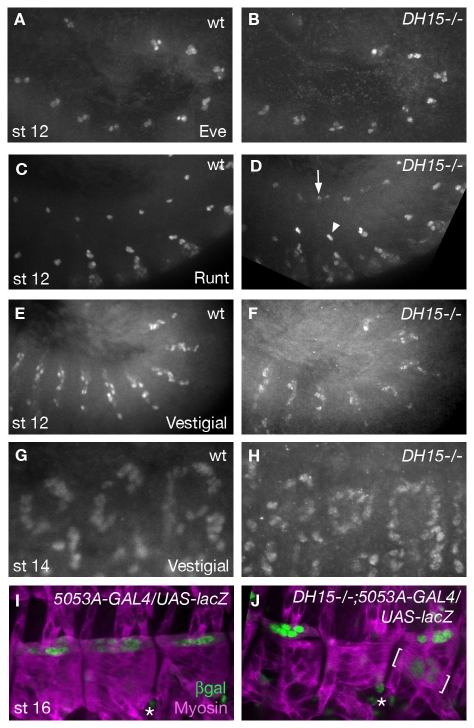
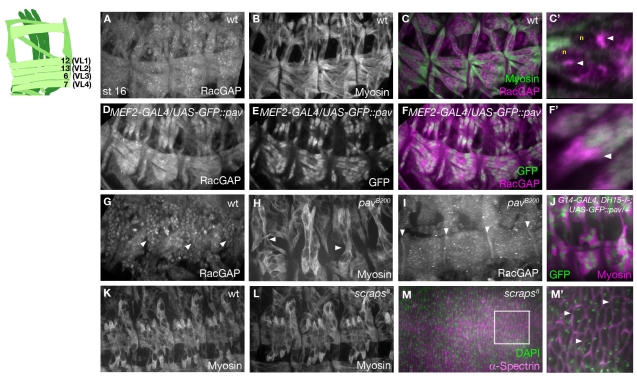
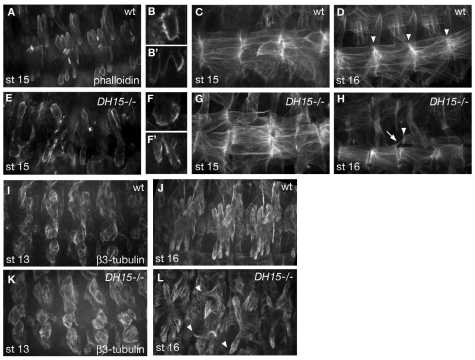
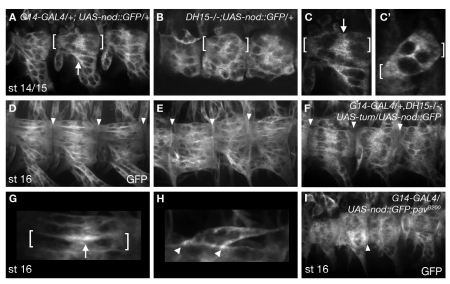
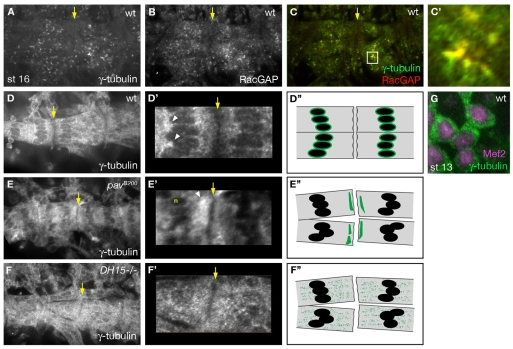
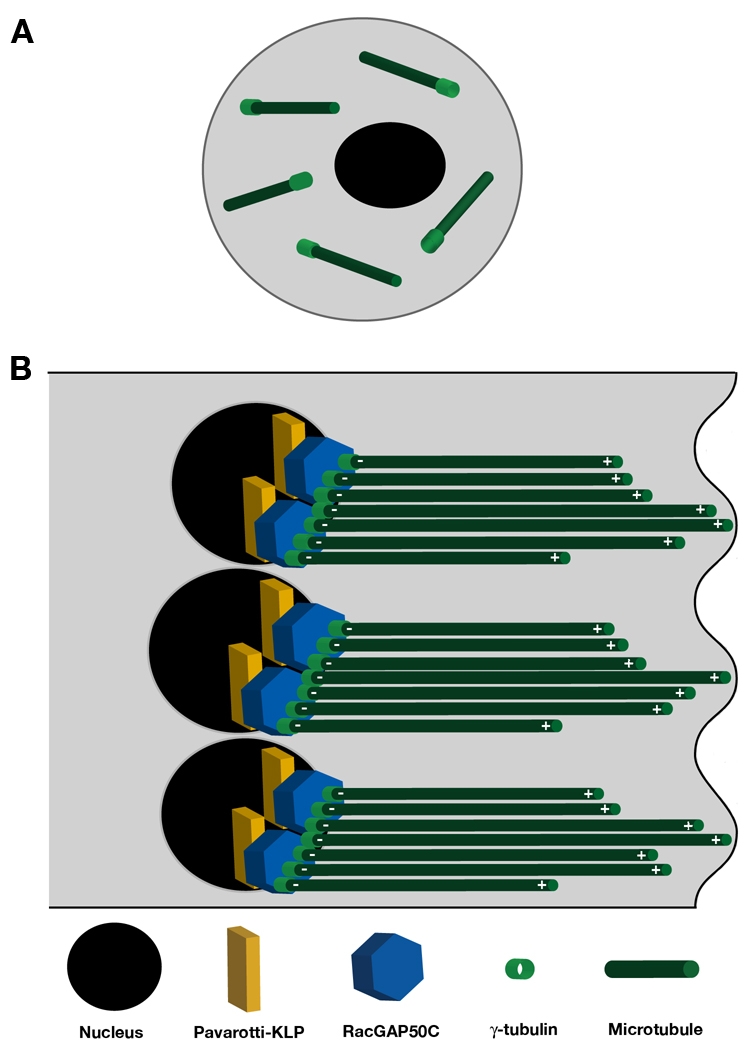
The microtubule (MT) cytoskeleton is reorganized during myogenesis as individual myoblasts fuse into multinucleated myotubes. Although this reorganization has long been observed in cell culture, these findings have not been validated during development, and proteins that regulate this process are largely unknown. We have identified a novel postmitotic function for the cytokinesis proteins RacGAP50C (Tumbleweed) and Pavarotti as essential regulators of MT organization during Drosophila myogenesis. We show that the localization of the MT nucleator gamma-tubulin changes from diffuse cytoplasmic staining in mononucleated myoblasts to discrete cytoplasmic puncta at the nuclear periphery in multinucleated myoblasts, and that this change in localization depends on RacGAP50C. RacGAP50C and gamma-tubulin colocalize at perinuclear sites in myotubes, and in RacGAP50C mutants gamma-tubulin remains dispersed throughout the cytoplasm. Furthermore, we show that the mislocalization of RacGAP50C in pavarotti mutants is sufficient to redistribute gamma-tubulin to the muscle fiber ends. Finally, myotubes in RacGAP50C mutants have MTs with non-uniform polarity, resulting in multiple guidance errors. Taken together, these findings provide strong evidence that the reorganization of the MT network that has been observed in vitro plays an important role in myotube extension and muscle patterning in vivo, and also identify two molecules crucial for this process.
Figures
Similar articles
-
Tum/RacGAP50C provides a critical link between anaphase microtubules and the assembly of the contractile ring in Drosophila melanogaster.J Cell Sci. 2005 Nov 15;118(Pt 22):5381-92. doi: 10.1242/jcs.02652. J Cell Sci. 2005. PMID: 16280552
-
A RhoGEF and Rho family GTPase-activating protein complex links the contractile ring to cortical microtubules at the onset of cytokinesis.Dev Cell. 2003 Jan;4(1):29-39. doi: 10.1016/s1534-5807(02)00402-1. Dev Cell. 2003. PMID: 12530961
-
Girds 'n' cleeks o' cytokinesis: microtubule sticks and contractile hoops in cell division.Biochem Soc Trans. 2008 Jun;36(Pt 3):400-4. doi: 10.1042/BST0360400. Biochem Soc Trans. 2008. PMID: 18481968
-
Targeting of γ-tubulin complexes to microtubule organizing centers: conservation and divergence.Trends Cell Biol. 2015 May;25(5):296-307. doi: 10.1016/j.tcb.2014.12.002. Epub 2014 Dec 24. Trends Cell Biol. 2015. PMID: 25544667 Review.
-
Acentrosomal microtubule nucleation in higher plants.Int Rev Cytol. 2002;220:257-89. doi: 10.1016/s0074-7696(02)20008-x. Int Rev Cytol. 2002. PMID: 12224551 Review.
Cited by
-
The Drosophila Ninein homologue Bsg25D cooperates with Ensconsin in myonuclear positioning.J Cell Biol. 2019 Feb 4;218(2):524-540. doi: 10.1083/jcb.201808176. Epub 2019 Jan 9. J Cell Biol. 2019. PMID: 30626718 Free PMC article.
-
A barley ROP GTPase ACTIVATING PROTEIN associates with microtubules and regulates entry of the barley powdery mildew fungus into leaf epidermal cells.Plant Cell. 2011 Jun;23(6):2422-39. doi: 10.1105/tpc.110.082131. Epub 2011 Jun 17. Plant Cell. 2011. PMID: 21685259 Free PMC article.
-
"Importin" signaling roles for import proteins: the function of Drosophila importin-7 (DIM-7) in muscle-tendon signaling.Cell Adh Migr. 2012 Jan-Feb;6(1):4-12. doi: 10.4161/cam.19774. Cell Adh Migr. 2012. PMID: 22647935 Free PMC article.
-
Dysferlin interacts with histone deacetylase 6 and increases alpha-tubulin acetylation.PLoS One. 2011;6(12):e28563. doi: 10.1371/journal.pone.0028563. Epub 2011 Dec 8. PLoS One. 2011. Retraction in: PLoS One. 2018 Jan 29;13(1):e0192239. doi: 10.1371/journal.pone.0192239. PMID: 22174839 Free PMC article. Retracted.
-
Microtubule Organization in Striated Muscle Cells.Cells. 2020 Jun 3;9(6):1395. doi: 10.3390/cells9061395. Cells. 2020. PMID: 32503326 Free PMC article. Review.
References
-
- Bartolini, F. and Gundersen, G. G. (2006). Generation of noncentrosomal microtubule arrays. J. Cell Sci. 119, 4155-4163. - PubMed
-
- Bate, M. (1990). The embryonic development of larval muscles in Drosophila. Development 110, 791-804. - PubMed
-
- Bate, M. and Rushton, E. (1993). Myogenesis and muscle patterning in Drosophila. CR Acad. Sci. III 316, 1047-1061. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
