

Review
doi: 10.1146/annurev.biochem.78.070507.135656.
The structural and functional diversity of metabolite-binding riboswitches
Affiliations
- PMID: 19298181
- PMCID: PMC5325118
- DOI: 10.1146/annurev.biochem.78.070507.135656
Item in Clipboard
Review
The structural and functional diversity of metabolite-binding riboswitches
Annu Rev Biochem.
2009.
Abstract
The cellular concentrations of certain metabolites are assiduously monitored to achieve appropriate levels of gene expression. Although proteins have long been known to act as sensors in this capacity, metabolite-binding RNAs, or riboswitches, also play an important role. More than 20 distinct classes of riboswitches have been identified to date, and insights to the molecular recognition strategies of a significant subset of these have been provided by detailed structural studies. This diverse set of metabolite-sensing RNAs is found to exploit a variety of distinct mechanisms to regulate genes that are fundamental to metabolism.
Figures
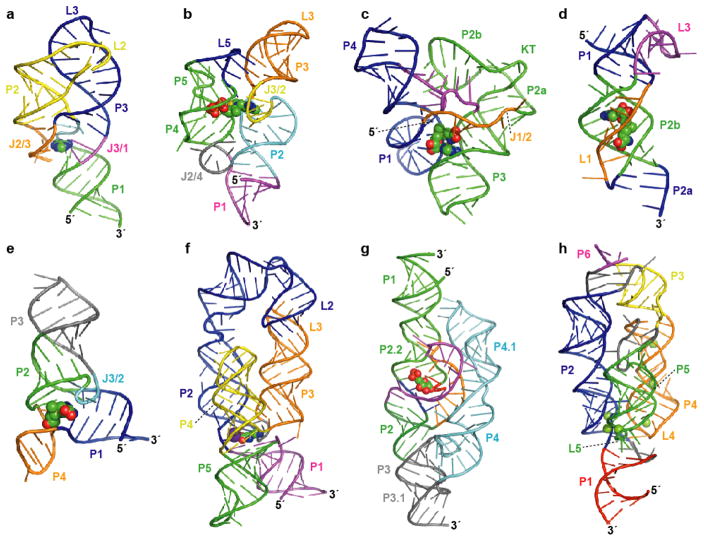
Atomic resolution structures for representatives of eight riboswitch aptamer classes. Aptamer structure models are shown as ribbon diagrams and ligands are represented as spheres. Pairing elements and joining regions are indicated. (a) The purine riboswitch aptamer (PDB ID 1Y26, 1Y27, 1U8D) (22, 23). (b) The TPP riboswitch aptamer (PDB ID 2GDI, 2HOJ, 2CKY) (–34). (c) The SAM-I riboswitch aptamer (PDB ID 2GIS) (44); KT, kink-turn. (d) The SAM-II riboswitch aptamer (PDB ID 2QWY) (45). (e) The SAM-III/SMK riboswitch aptamer (PDB ID 3E5C) (46). (f) The lysine riboswitch aptamer (PDB ID 3D0U, 3DIL) (56, 57); a specifically bound K+ ion (57) is depicted as a purple sphere. (g) The GlcN6P-responsive glmS ribozyme (PDB ID 2HO7, 2N74) (65, 66). (h) The Mg2+-responsive M-box riboswitch aptamer (PDB ID 2QBZ) (75); six crystallographically observed Mg2+ ions are shown.
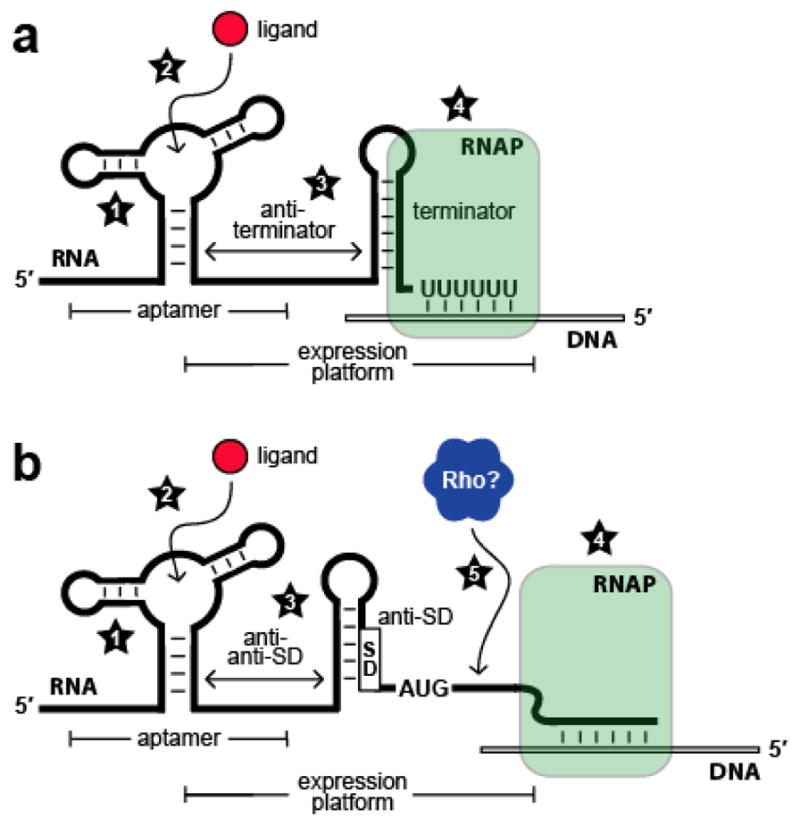
Kinetic and thermodynamic factors that influence riboswitch function. (a) Schematic representation of a riboswitch that represses gene expression upon ligand binding by controlling transcription termination. Factors that affect riboswitch function include (1) rate constants for aptamer folding and unfolding; (2) rate constants for ligand association and dissociation; (3) rate constants for expression platform folding and unfolding; (4) speed of transcription elongation by RNA polymerase (RNAP). (b) Schematic representation of a riboswitch that represses gene expression upon ligand binding by controlling translation initiation. Numbers 1 through 4 are as described in (a); (5) speed of Rho-dependent transcription termination.
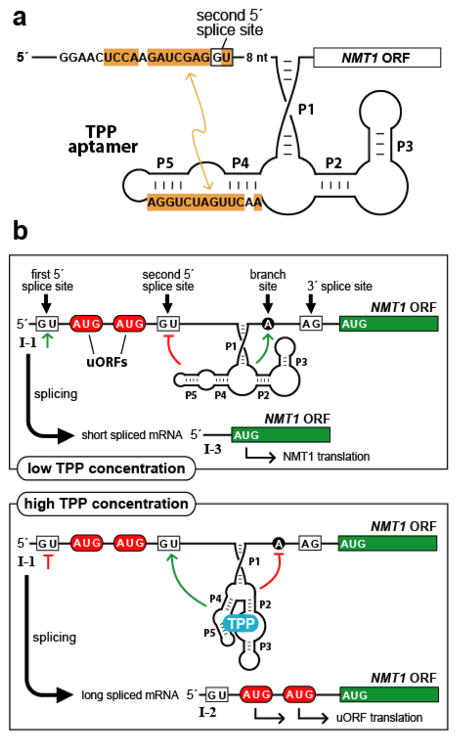
Gene control by a eukaryotic TPP riboswitch. (a) Secondary structure model of the TPP aptamer residing in the 5′ UTR of Neurospora crassa NMT1. Sequences of the aptamer P4/P5 domain are complementary (orange shading) to a region adjoining a key splice site, illustrating one way in which the availability of TPP influences splice site selection. Similar types of base-pairing interactions between aptamer domains and expression platforms are exploited by other eukaryotic TPP riboswitches. (b) Control of alternative splicing by TPP riboswitches regulates NMT1 gene expression in fungi. The main features are depicted of the riboswitch from N. crassa NMT1, including splice sites, pairing elements of the TPP aptamer (P1 through P5), and uORFs that compete with translation of the primary ORF to reduce gene expression. Green lines and red lines refer to splicing determinants that are activated or inhibited, respectively, depending on the occupancy state of the aptamer domain.
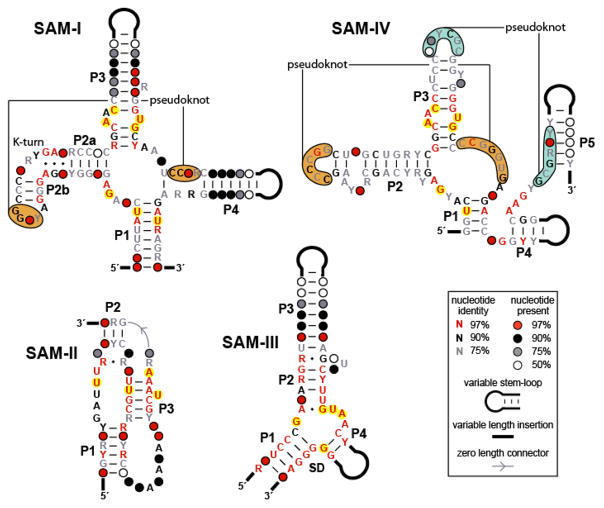
Conserved sequences and secondary structure models corresponding to different classes of SAM riboswitch aptamers. The SAM-I, SAM-II, and SAM-III classes have distinct architectures, while the SAM-IV aptamer is related in sequence and secondary structure to the SAM-I class. Nucleotides highlighted in yellow are observed to contact the SAM ligand directly (–46). Note that, with only a single exception, the SAM-IV aptamer consensus structure contains these same conserved nucleotides in analogous positions, suggesting that the binding pockets are highly similar. Distinct but positionally analogous pseudoknots in the SAM-I and SAM-IV motifs are indicated by orange shading, while an additional predicted pseudoknot unique to the SAM-IV class is denoted by blue shading.
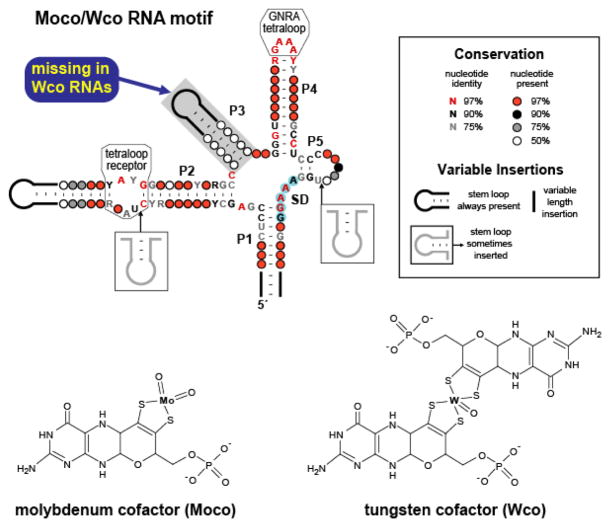
Conserved sequences and secondary structure models corresponding to Moco and Wco RNAs. These RNAs exhibit characteristics of riboswitches, including a complex aptamer-like structure and gene control function. Many representatives reside immediately upstream of an AUG start codon and are predicted to function by sequestering the SD sequence (blue shading). The phylogenetic distributions and specific gene associations of these motifs suggest that RNAs containing the P3 element recognize Moco, while those lacking this stem bind the related coenzyme Wco.
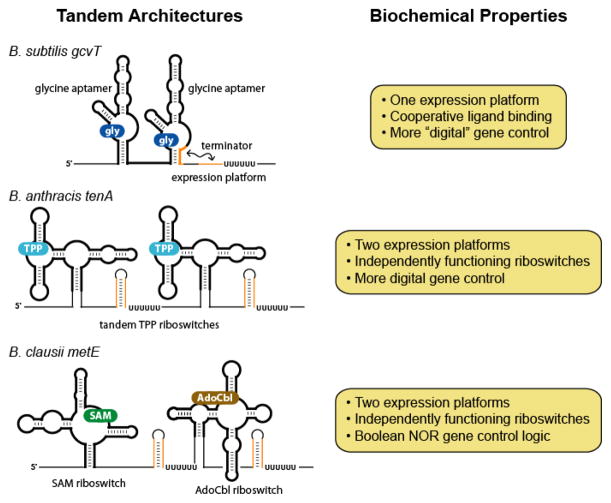
Tandem arrangements of aptamers and riboswitches. The schematic diagrams illustrate some of the distinct ways in which riboswitches or their component aptamers can be combined to effect more sophisticated genetic control.
References
-
- Steitz TA, Moore PB. RNA, the first macromolecular catalyst: the ribosome is a ribozyme. Trends Biochem Sci. 2003;28:411–8. - PubMed
-
- Valadkhan S, Manley JL. Splicing-related catalysis by protein-free snRNAs. Nature. 2001;413:701–7. - PubMed
-
- Doudna JA, Batey RT. Structural insights into the signal recognition particle. Annu Rev Biochem. 2004;73:539–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
