

Estrogen-induced plasticity from cells to circuits: predictions for cognitive function
- PMID: 19299024
- PMCID: PMC3167490
- DOI: 10.1016/j.tips.2008.12.006
Estrogen-induced plasticity from cells to circuits: predictions for cognitive function
Abstract
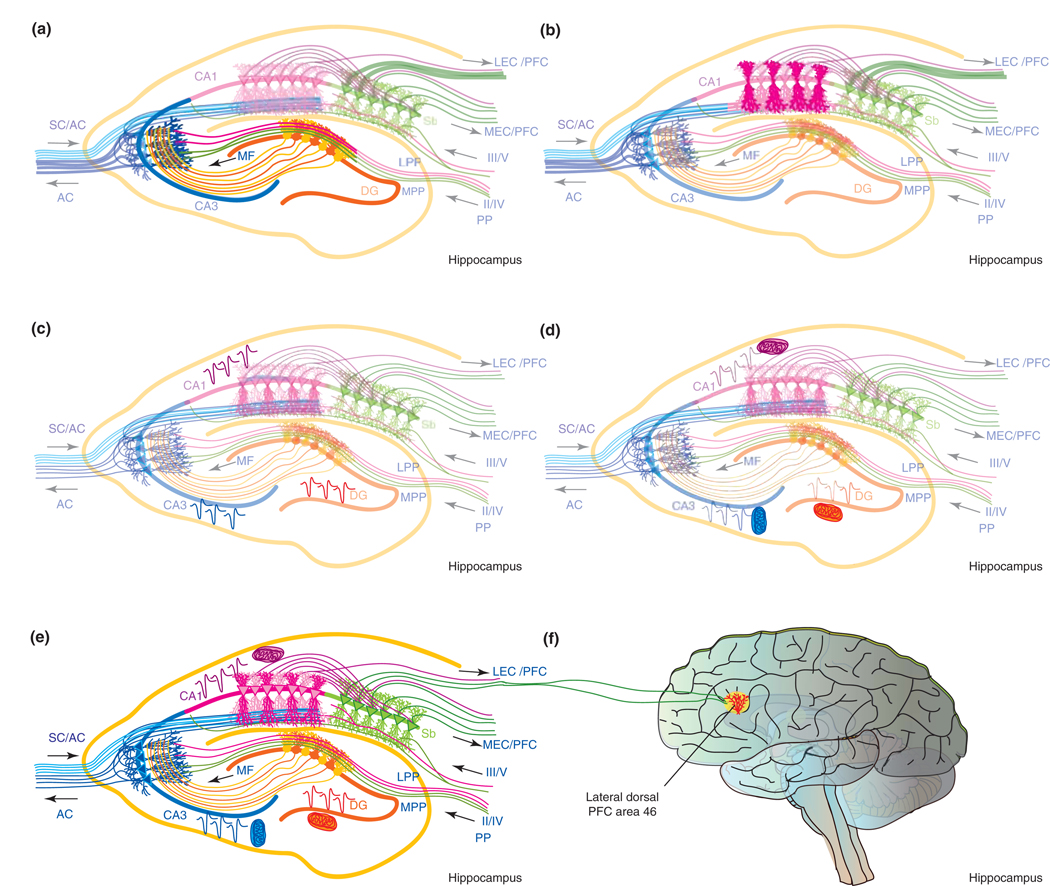
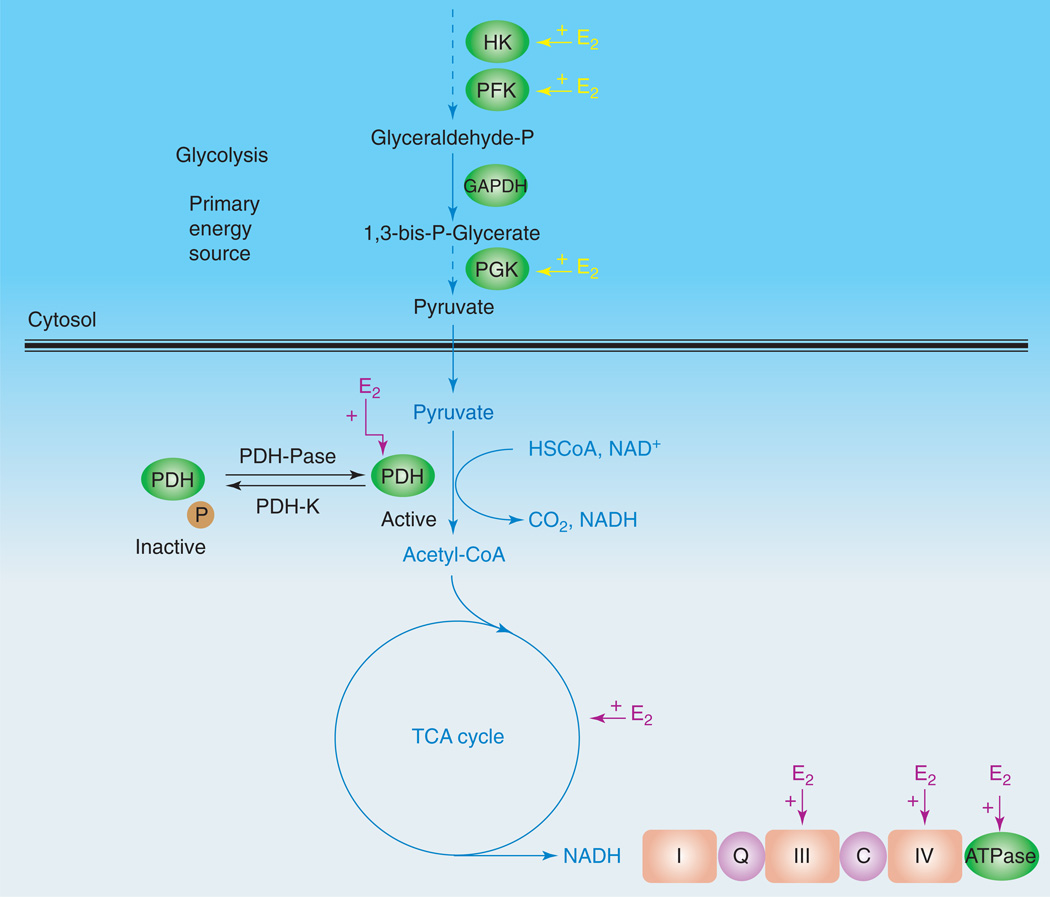
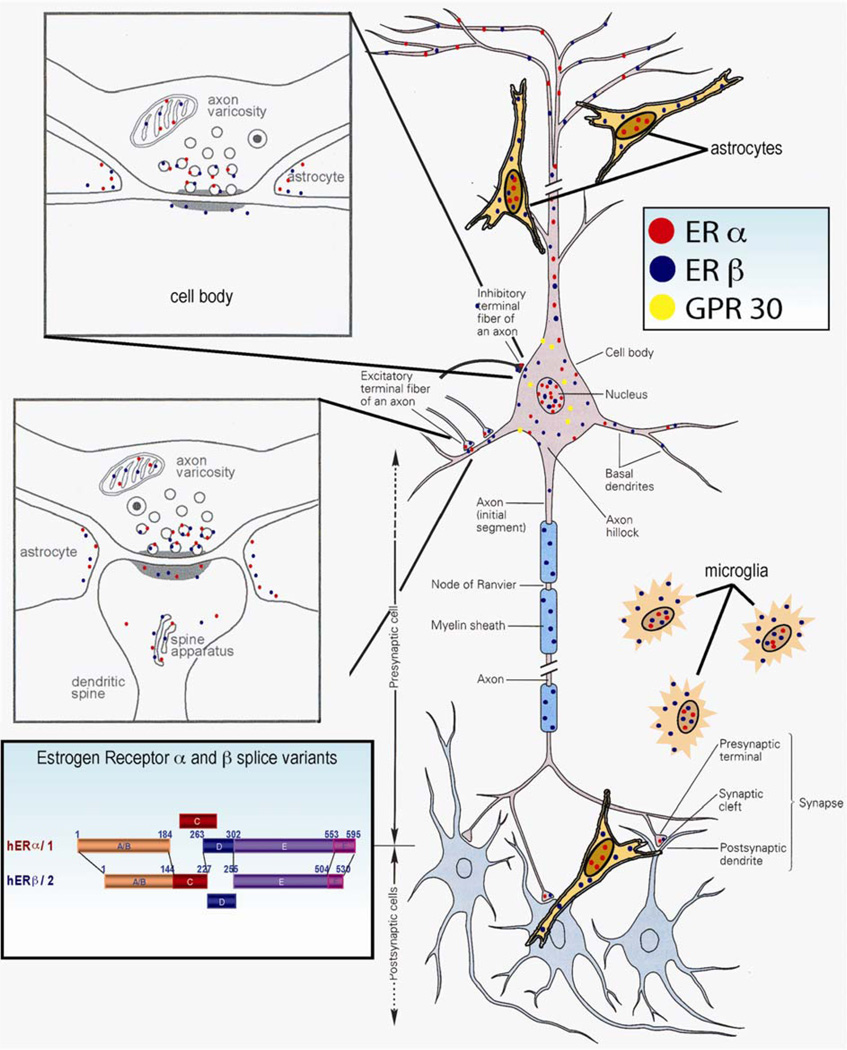
Controversy regarding estrogen action in the brain remains at the forefront of basic, translational and clinical science for women's health. Here, I provide an integrative analysis of estrogen-inducible plasticity and posit it as a strategy for predicting cognitive domains affected by estrogen in addition to sources of variability. Estrogen enhancement of plasticity is evidenced by increases in neurogenesis, neural network connectivity and synaptic transmission. In parallel, estrogen increases glucose transport, aerobic glycolysis and mitochondrial function to provide the ATP necessary to sustain increased energetic demand. The pattern of plasticity predicts that estrogen would preferentially affect cognitive tasks of greater complexity, temporal demand and associative challenge. Thus, estrogen deprivation should be associated with decrements in these functions. Estrogen regulation of plasticity and bioenergetics provides a framework for predicting estrogen-dependent cognitive functions while also identifying sources of variability and potential biomarkers for identifying women appropriate for hormone therapy.
Figures
References
-
- Brinton RD. Investigative models for determining hormone therapy-induced outcomes in brain: evidence in support of a healthy cell bias of estrogen action. Ann. N. Y. Acad. Sci. 2005;1052:57–74. - PubMed
-
- Hogervorst E, et al. The nature of the effect of female gonadal hormone replacement therapy on cognitive function in post-menopausal women: a meta-analysis. Neuroscience. 2000;101:485–512. - PubMed
-
- Sherwin BB, Henry JF. Brain aging modulates the neuroprotective effects of estrogen on selective aspects of cognition in women: a critical review. Front. Neuroendocrinol. 2008;29:88–113. - PubMed
-
- Zhao C, et al. Mechanisms and functional implications of adult neurogenesis. Cell. 2008;132:645–660. - PubMed
-
- Tanapat P, et al. Ovarian steroids influence cell proliferation in the dentate gyrus of the adult female rat in a dose- and time-dependent manner. J. Comp. Neurol. 2005;481:252–265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
