

Perilipin is present in islets of Langerhans and protects against lipotoxicity when overexpressed in the beta-cell line INS-1
- PMID: 19299455
- PMCID: PMC2703509
- DOI: 10.1210/en.2008-0913
Perilipin is present in islets of Langerhans and protects against lipotoxicity when overexpressed in the beta-cell line INS-1
Abstract
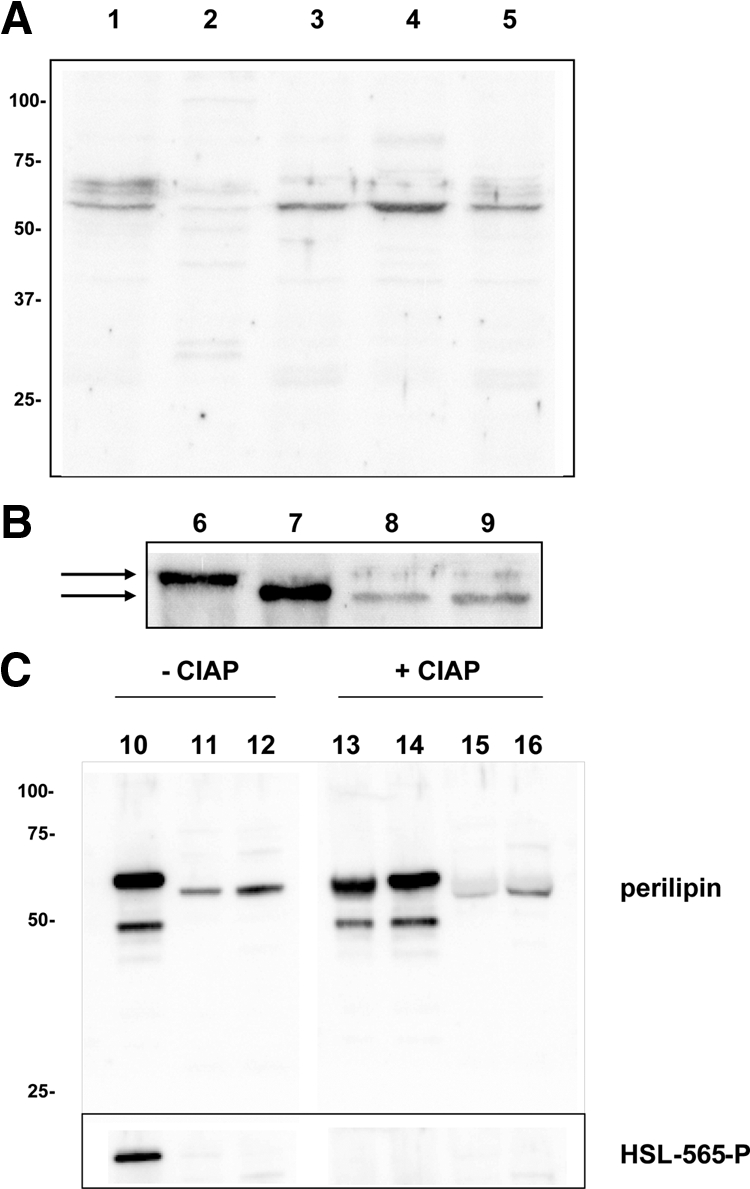
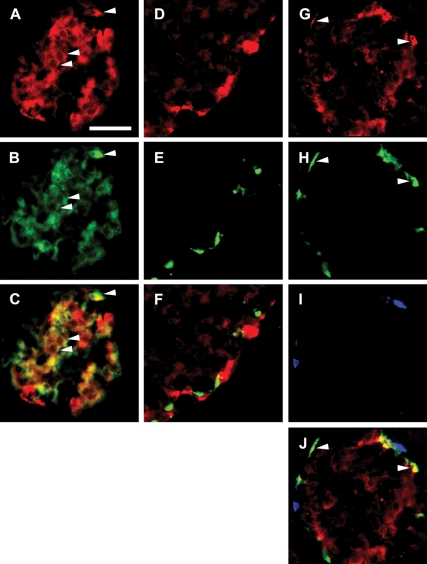
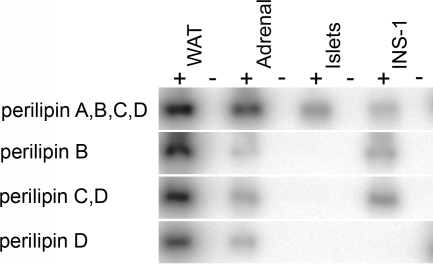
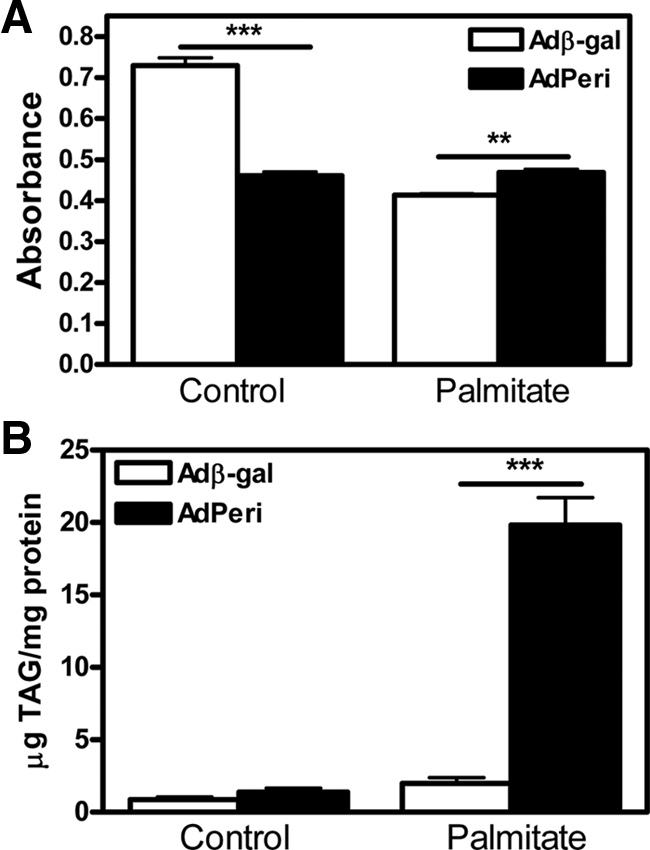
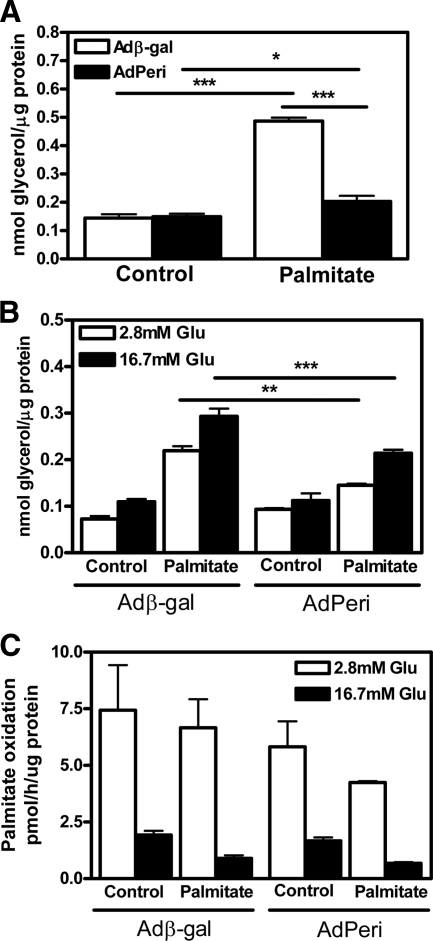
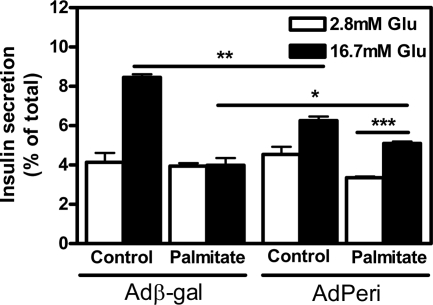
Lipids have been shown to play a dual role in pancreatic beta-cells: a lipid-derived signal appears to be necessary for glucose-stimulated insulin secretion, whereas lipid accumulation causes impaired insulin secretion and apoptosis. The ability of the protein perilipin to regulate lipolysis prompted an investigation of the presence of perilipin in the islets of Langerhans. In this study evidence is presented for perilipin expression in rat, mouse, and human islets of Langerhans as well as the rat clonal beta-cell line INS-1. In rat and mouse islets, perilipin was verified to be present in beta-cells. To examine whether the development of lipotoxicity could be prevented by manipulating the conditions for lipid storage in the beta-cell, INS-1 cells with adenoviral-mediated overexpression of perilipin were exposed to lipotoxic conditions for 72 h. In cells exposed to palmitate, perilipin overexpression caused increased accumulation of triacylglycerols and decreased lipolysis compared with control cells. Whereas glucose-stimulated insulin secretion was retained after palmitate exposure in cells overexpressing perilipin, it was completely abolished in control beta-cells. Thus, overexpression of perilipin appears to confer protection against the development of beta-cell dysfunction after prolonged exposure to palmitate by promoting lipid storage and limiting lipolysis.
Figures

References
-
- Henquin JC 2000 Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 49:1751–1760 - PubMed
-
- Mulder H, Yang S, Winzell MS, Holm C, Ahrén B 2004 Inhibition of lipase activity and lipolysis in rat islets reduces insulin secretion. Diabetes 53:122–128 - PubMed
-
- Koyama K, Chen G, Wang MY, Lee Y, Shimabukuro M, Newgard CB, Unger RH 1997 β-Cell function in normal rats made chronically hyperleptinemic by adenovirus-leptin gene therapy. Diabetes 46:1276–1280 - PubMed
-
- Masiello P, Novelli M, Bombara M, Fierabracci V, Vittorini S, Prentki M, Bergamini E 2002 The antilipolytic agent 3,5-dimethylpyrazole inhibits insulin release in response to both nutrient secretagogues and cyclic adenosine monophosphate agonists in isolated rat islets. Metabolism 51:110–114 - PubMed
