

A Golgi-associated protein 4.1B variant is required for assimilation of proteins in the membrane
- PMID: 19299464
- PMCID: PMC2714437
- DOI: 10.1242/jcs.039644
A Golgi-associated protein 4.1B variant is required for assimilation of proteins in the membrane
Erratum in
-
Correction: A Golgi-associated protein 4.1B variant is required for assimilation of proteins in the membrane (doi:10.1242/jcs.039644).J Cell Sci. 2019 Oct 21;132(20):jcs240002. doi: 10.1242/jcs.240002. J Cell Sci. 2019. PMID: 31636159 Free PMC article. No abstract available.
Expression of concern in
-
Expression of Concern: A Golgi-associated protein 4.1B variant is required for assimilation of proteins in the membrane (doi:10.1242/jcs.039644).J Cell Sci. 2019 May 2;132(9):jcs233080. doi: 10.1242/jcs.233080. J Cell Sci. 2019. PMID: 31048547 Free PMC article. No abstract available.
Abstract
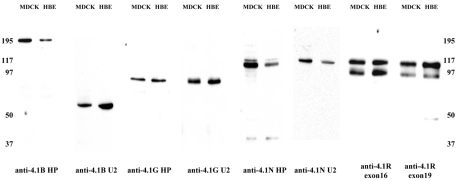
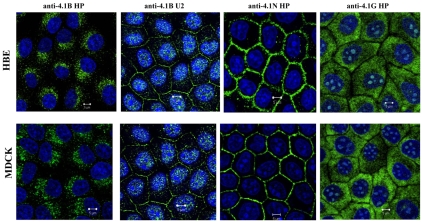
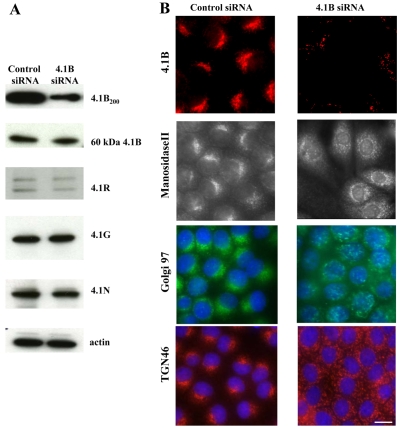
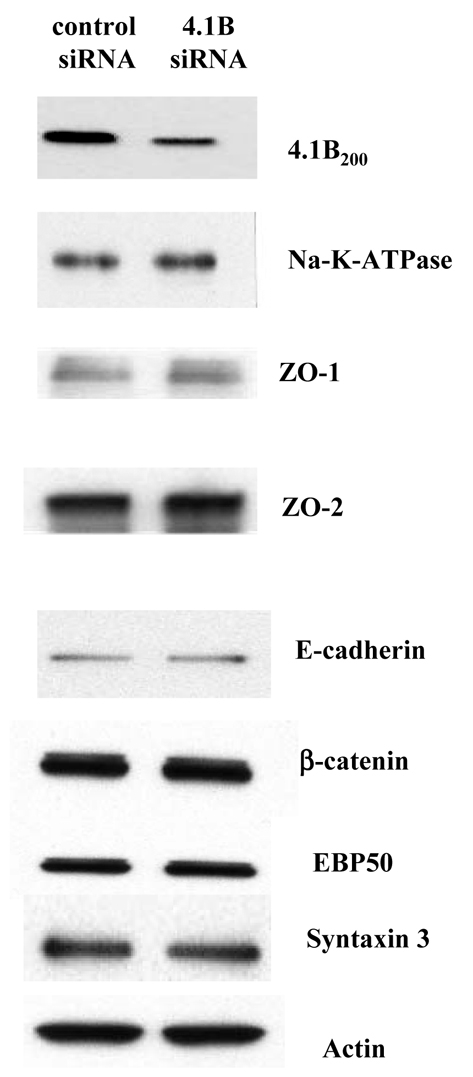
The archetypal membrane skeleton is that of the erythrocyte, consisting predominantly of spectrin, actin, ankyrin R and protein 4.1R. The presence in the Golgi of a membrane skeleton with a similar structure has been inferred, based on the identification of Golgi-associated spectrin and ankyrin. It has long been assumed that a Golgi-specific protein 4.1 must also exist, but it has not previously been found. We demonstrate here that a hitherto unknown form of protein 4.1, a 200 kDa 4.1B, is associated with the Golgi of Madin-Darby canine kidney (MDCK) and human bronchial epithelial (HBE) cells. This 4.1B variant behaves like a Golgi marker after treatment with Brefeldin A and during mitosis. Depletion of the protein in HBE cells by siRNA resulted in disruption of the Golgi structure and failure of Na(+)/K(+)-ATPase, ZO-1 and ZO-2 to migrate to the membrane. Thus, this newly identified Golgi-specific protein 4.1 appears to have an essential role in maintaining the structure of the Golgi and in assembly of a subset of membrane proteins.
Figures






References
-
- Aunis, D. and Bader, M. F. (1988). The cytoskeleton as a barrier to exocytosis in secretory cells. J. Exp. Biol. 139, 253-266. - PubMed
-
- Baines, A. J. and Bennett, V. (1985). Synapsin I is a spectrin-binding protein immunologically related to erythrocyte protein 4.1. Nature 315, 410-413. - PubMed
-
- Barr, F. A. and Short, B. (2003). Golgins in the structure and dynamics of the Golgi apparatus. Curr. Opin. Cell Biol. 15, 405-413. - PubMed
-
- Beck, K. A. (2005). Spectrins and the Golgi. Biochim. Biophys. Acta 1744, 374-382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
