

Characterization of a thermostable archaeal polynucleotide kinase homologous to human Clp1
- PMID: 19299550
- PMCID: PMC2673061
- DOI: 10.1261/rna.1492809
Characterization of a thermostable archaeal polynucleotide kinase homologous to human Clp1
Abstract
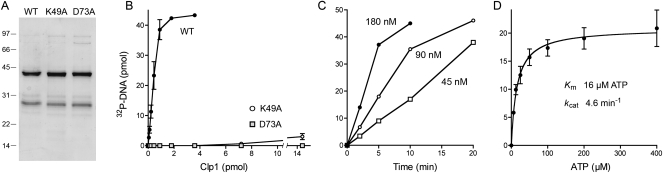
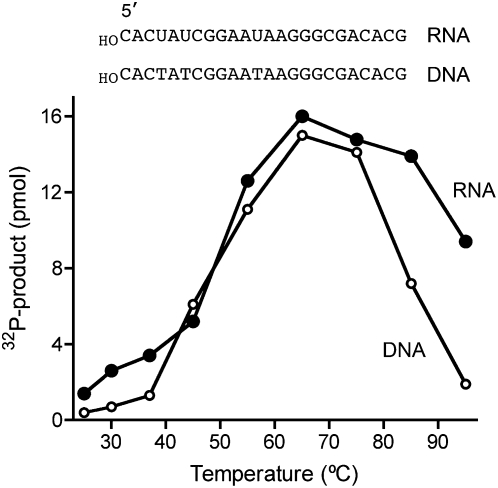
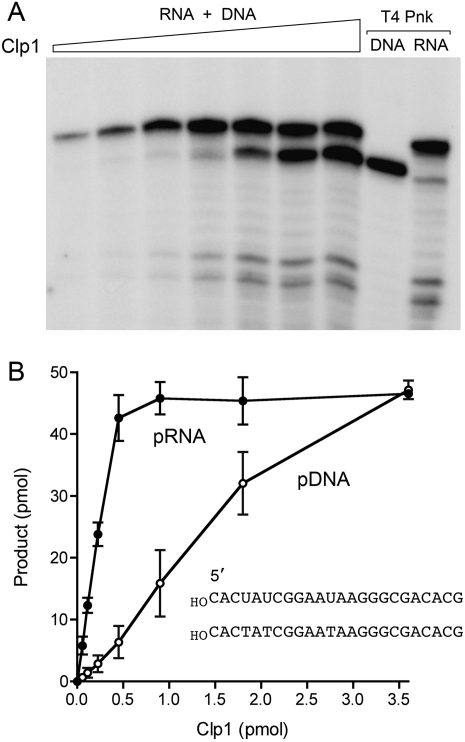
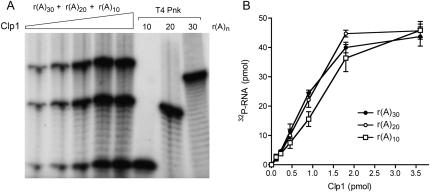
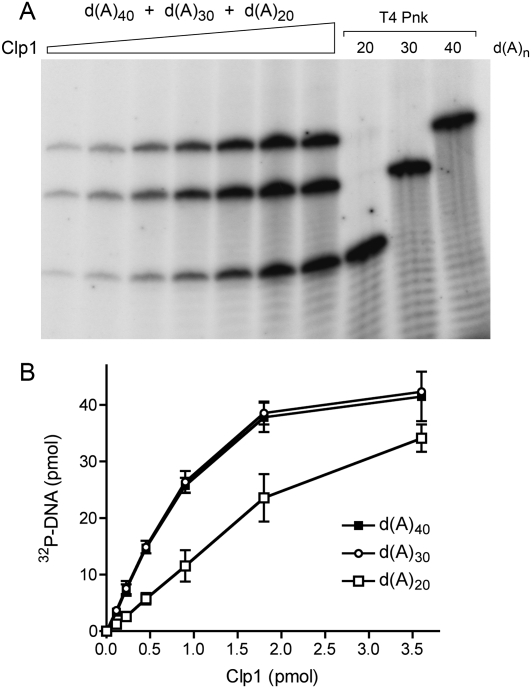
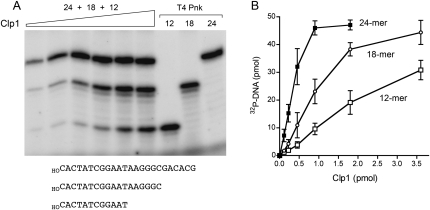
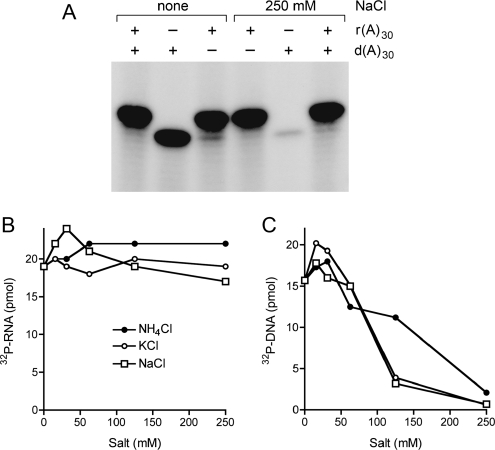
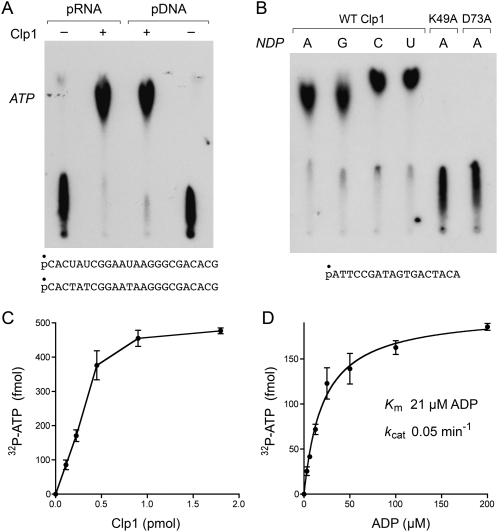
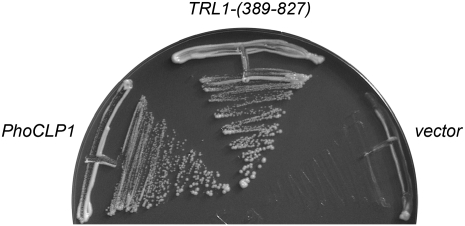
Clp1 proteins are essential components of the eukaryal mRNA 3' cleavage-polyadenylation machinery. Human Clp1 has an additional function as an RNA-specific 5'-OH polynucleotide kinase, which is implicated in RNA end healing. Yeast Clp1 has no kinase activity, although it binds ATP. Here we report that Clp1-like proteins are extant in archaea. Purification and characterization of Pyrococcus horikoshii Clp1 (PhoClp1) reveals it to be a thermostable 5'-OH polynucleotide kinase optimally active at 55 degrees C to 85 degrees C. PhoClp1 catalyzes transfer of the gamma phosphate from ATP (K (m) 16 microM) to either 5'-OH RNA or DNA ends, although it prefers RNA in a competitive situation. Increasing the monovalent salt concentration to 250 mM suppresses the DNA kinase without affecting RNA phosphorylation, suggesting that RNA is a likely substrate for this enzyme in vivo. Indeed, we show that expression of PhoClp1 in budding yeast can complement a lethal mutation in the 5'-OH RNA kinase module of tRNA ligase. PhoClp1 is a member of the P-loop phosphotransferase superfamily. Alanine mutations at the P-loop lysine (Lys49) and a conserved aspartate (Asp73) inactivate the kinase. Our studies fortify emerging evidence for an enzymatic RNA repair capacity in archaea and provide a new reagent for polynucleotide phosphorylation at high temperatures.
Figures

References
-
- Barrangou R., Fremauzx C., Deveau H., Richards M., Boyaval P., Moineau S., Romero D.A., Horvath P. CRISPR provides acquired resistance against viruses in prokaryotes. Science. 2007;315:1709–1712. - PubMed
-
- Bernstein N.K., Williams R.S., Rakovszky M.L., Cui D., Green R., Galicia S., Koch C.A., Cass C.E., Durocher D., Weinfeld M., et al. The molecular architecture of the mammalian DNA repair enzyme, polynucleotide kinase. Mol. Cell. 2005;17:657–670. - PubMed
-
- Blondal T., Hjorleifsdottir S., Aevarsson A., Fridjonsson O.H., Skirnisdottir S., Wheat J.O., Hermannsdottir A.G., Hreggvidsson G.O., Smith A.V., Kristjansson J.K. Characterization of a 5′-polynucleotide kinase/3′-phosphatase from bacteriophage RM378. J. Biol. Chem. 2005;280:5188–5194. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases