

Cooperative NCoR/SMRT interactions establish a corepressor-based strategy for integration of inflammatory and anti-inflammatory signaling pathways
- PMID: 19299558
- PMCID: PMC2661610
- DOI: 10.1101/gad.1773109
Cooperative NCoR/SMRT interactions establish a corepressor-based strategy for integration of inflammatory and anti-inflammatory signaling pathways
Abstract
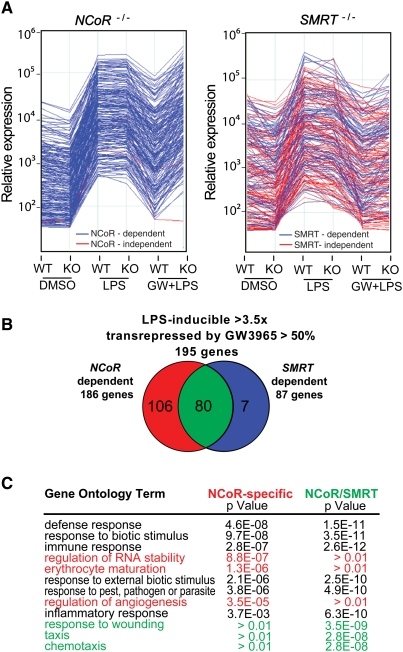
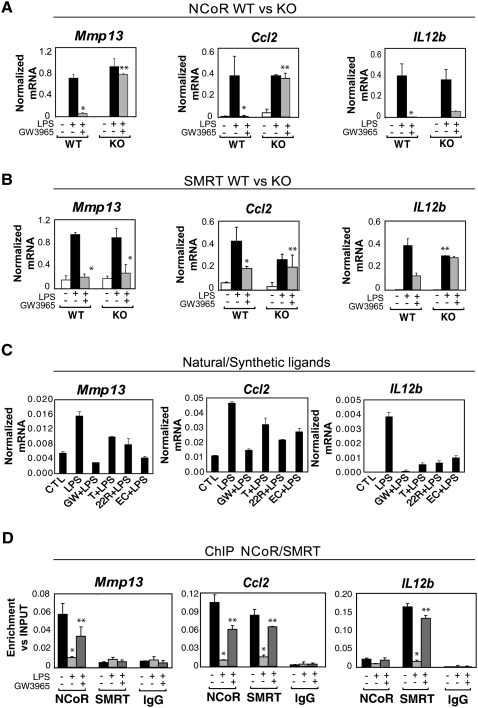
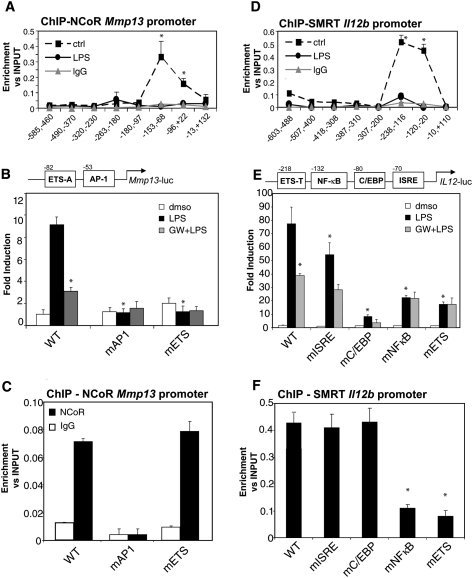
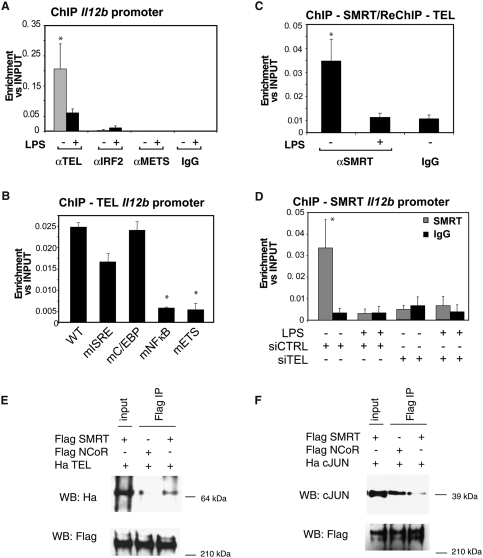
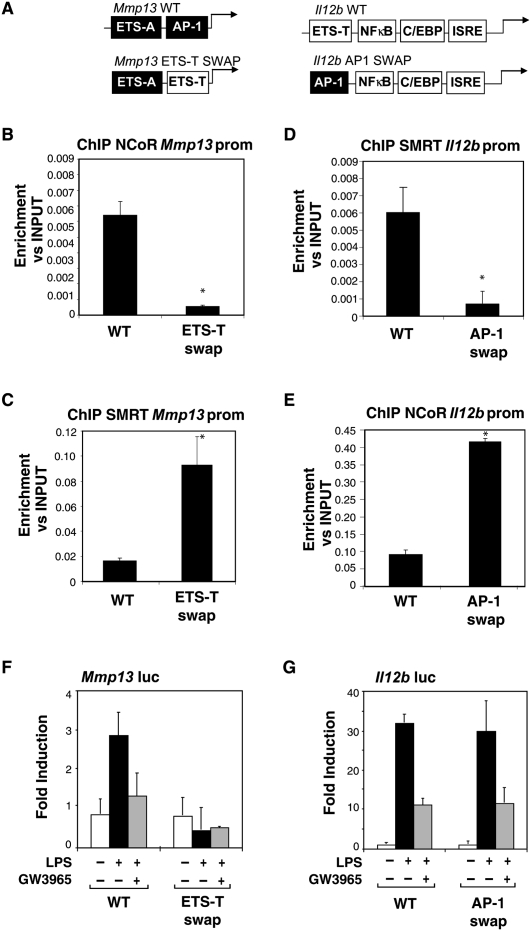
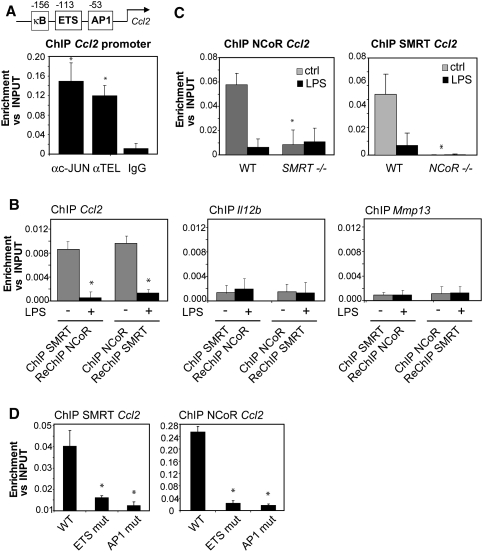
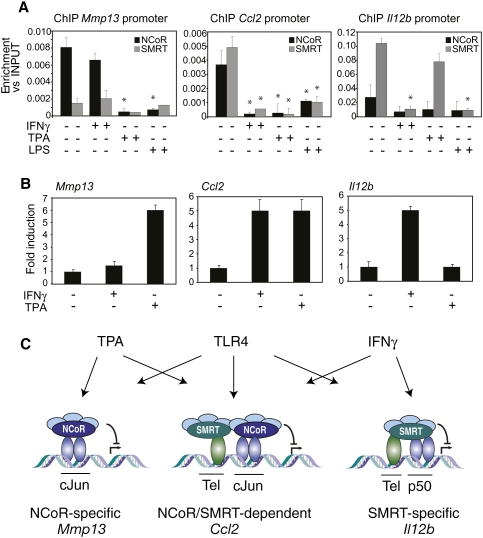
Innate immune responses to bacterial or viral infection require rapid transition of large cohorts of inflammatory response genes from poised/repressed to actively transcribed states, but the underlying repression/derepression mechanisms remain poorly understood. Here, we report that, while the nuclear receptor corepressor (NCoR) and silencing mediator of retinoic acid and thyroid hormone receptor (SMRT) corepressors establish repression checkpoints on broad sets of inflammatory response genes in macrophages and are required for nearly all of the transrepression activities of liver X receptors (LXRs), they can be selectively recruited via c-Jun or the Ets repressor Tel, respectively, establishing NCoR-specific, SMRT-specific, and NCoR/SMRT-dependent promoters. Unexpectedly, the binding of NCoR and SMRT to NCoR/SMRT-dependent promoters is frequently mutually dependent, establishing a requirement for both proteins for LXR transrepression and enabling inflammatory signaling pathways that selectively target NCoR or SMRT to also derepress/activate NCoR/SMRT-dependent genes. These findings reveal a combinatorial, corepressor-based strategy for integration of inflammatory and anti-inflammatory signals that play essential roles in immunity and homeostasis.
Figures
References
-
- Akira S., Takeda K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004;4:499–511. - PubMed
-
- Angel P., Imagawa M., Chiu R., Stein B., Imbra R.J., Rahmsdorf H.J., Jonat C., Herrlich P., Karin M. Phorbol ester-inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell. 1987;49:729–739. - PubMed
-
- Baek S.H., Ohgi K.A., Rose D.W., Koo E.H., Glass C.K., Rosenfeld M.G. Exchange of N-CoR corepressor and Tip60 coactivator complexes links gene expression by NF-κB and β-amyloid precursor protein. Cell. 2002;110:55–67. - PubMed
-
- Blaschke F., Takata Y., Caglayan E., Collins A., Tontonoz P., Hsueh W.A., Tangirala R.K. A nuclear receptor corepressor-dependent pathway mediates suppression of cytokine-induced C-reactive protein gene expression by liver X receptor. Circ. Res. 2006;99:e88–e99. doi: 10.1161/01.RES.0000252878.34269.06. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DK3949/DK/NIDDK NIH HHS/United States
- U54 GM069338/GM/NIGMS NIH HHS/United States
- R01 CA052599/CA/NCI NIH HHS/United States
- P30 DK063491/DK/NIDDK NIH HHS/United States
- DK063491/DK/NIDDK NIH HHS/United States
- DK074868/DK/NIDDK NIH HHS/United States
- CA052599/CA/NCI NIH HHS/United States
- GM069338/GM/NIGMS NIH HHS/United States
- P01 HL088093/HL/NHLBI NIH HHS/United States
- P01 DK074868/DK/NIDDK NIH HHS/United States
- HL088093/HL/NHLBI NIH HHS/United States
- HL065445/HL/NHLBI NIH HHS/United States
- R01 HL065445/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous