

Reduced c-myc expression levels limit follicular mature B cell cycling in response to TLR signals
- PMID: 19299704
- PMCID: PMC3441183
- DOI: 10.4049/jimmunol.0802961
Reduced c-myc expression levels limit follicular mature B cell cycling in response to TLR signals
Abstract
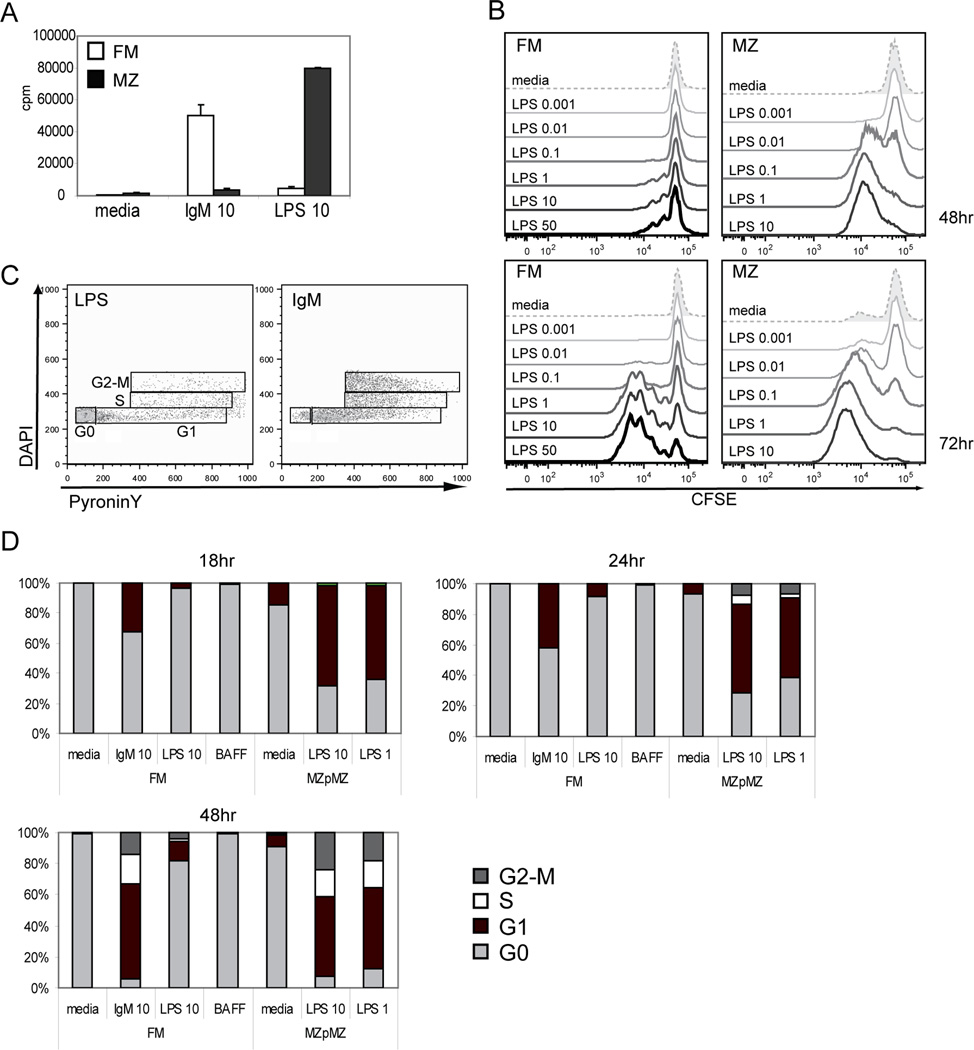
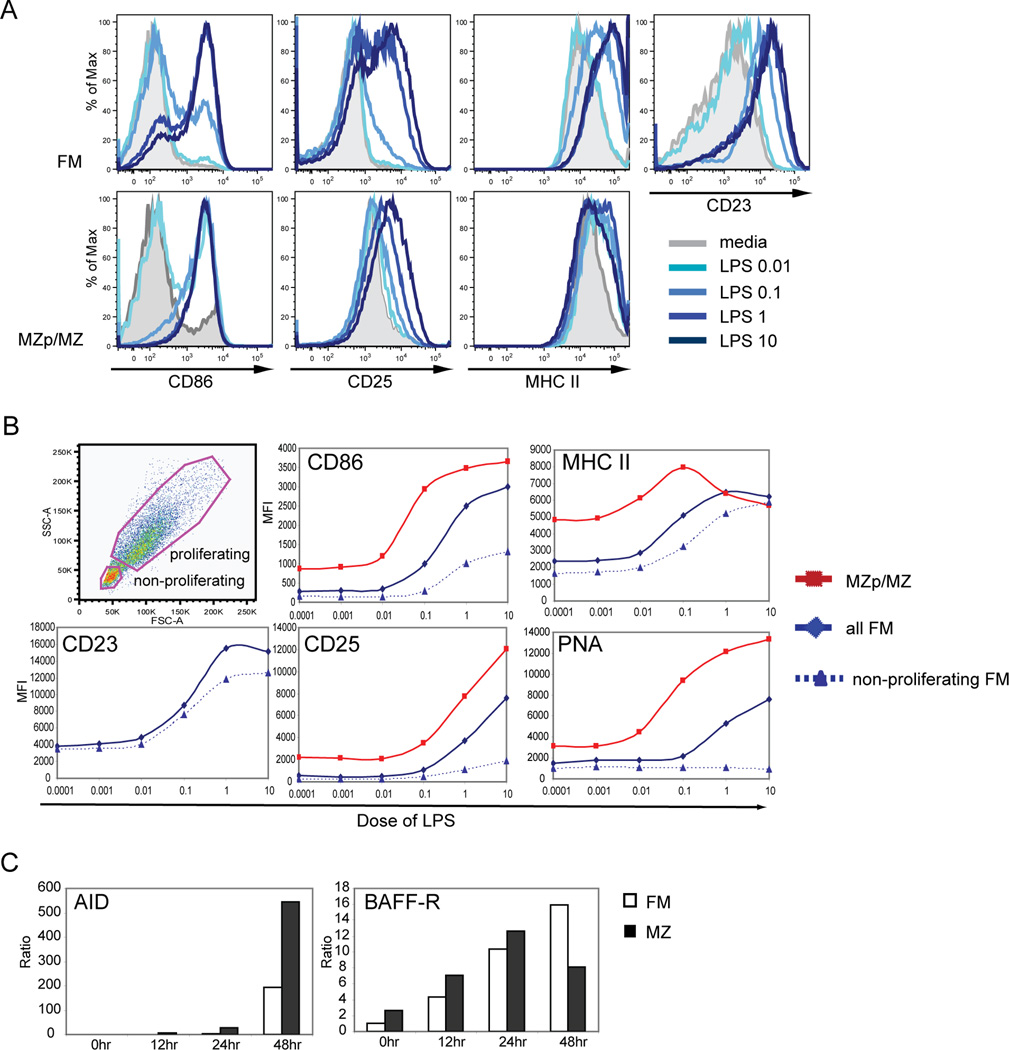
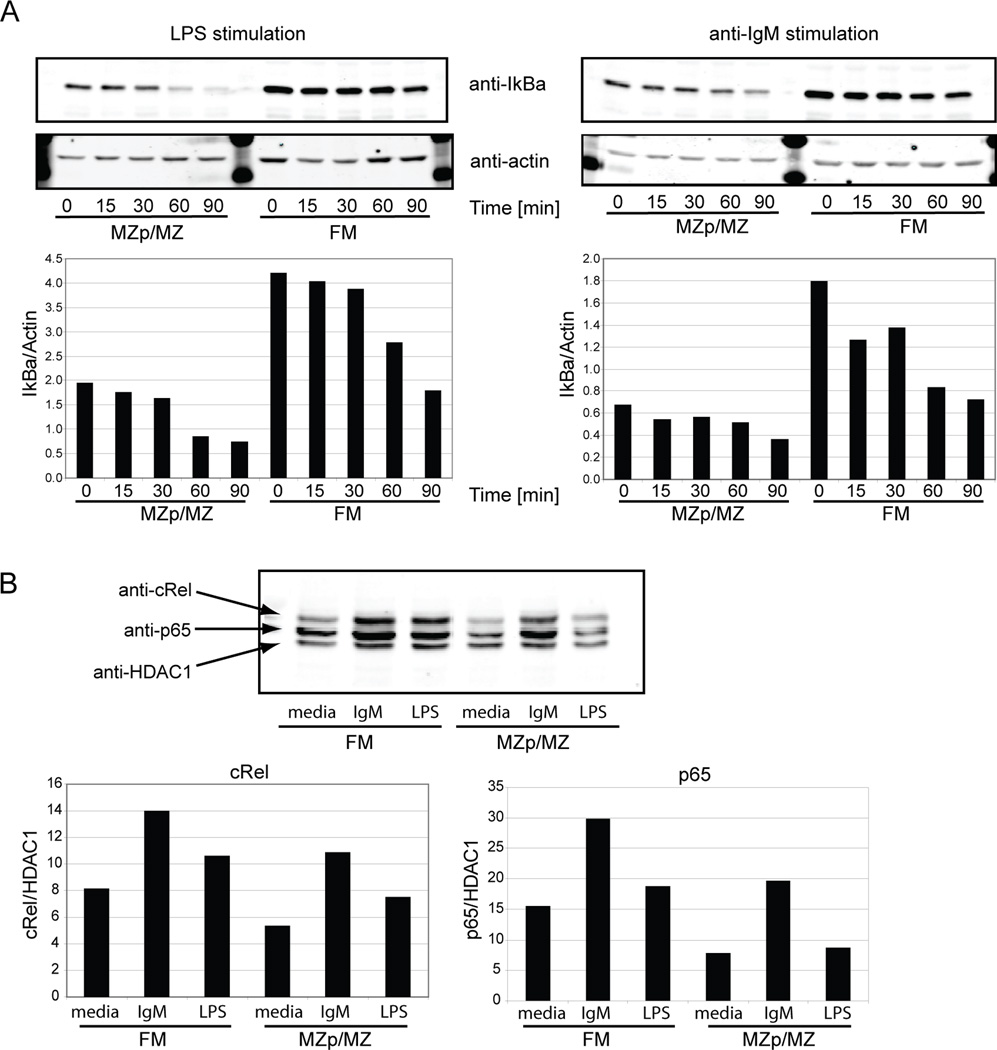
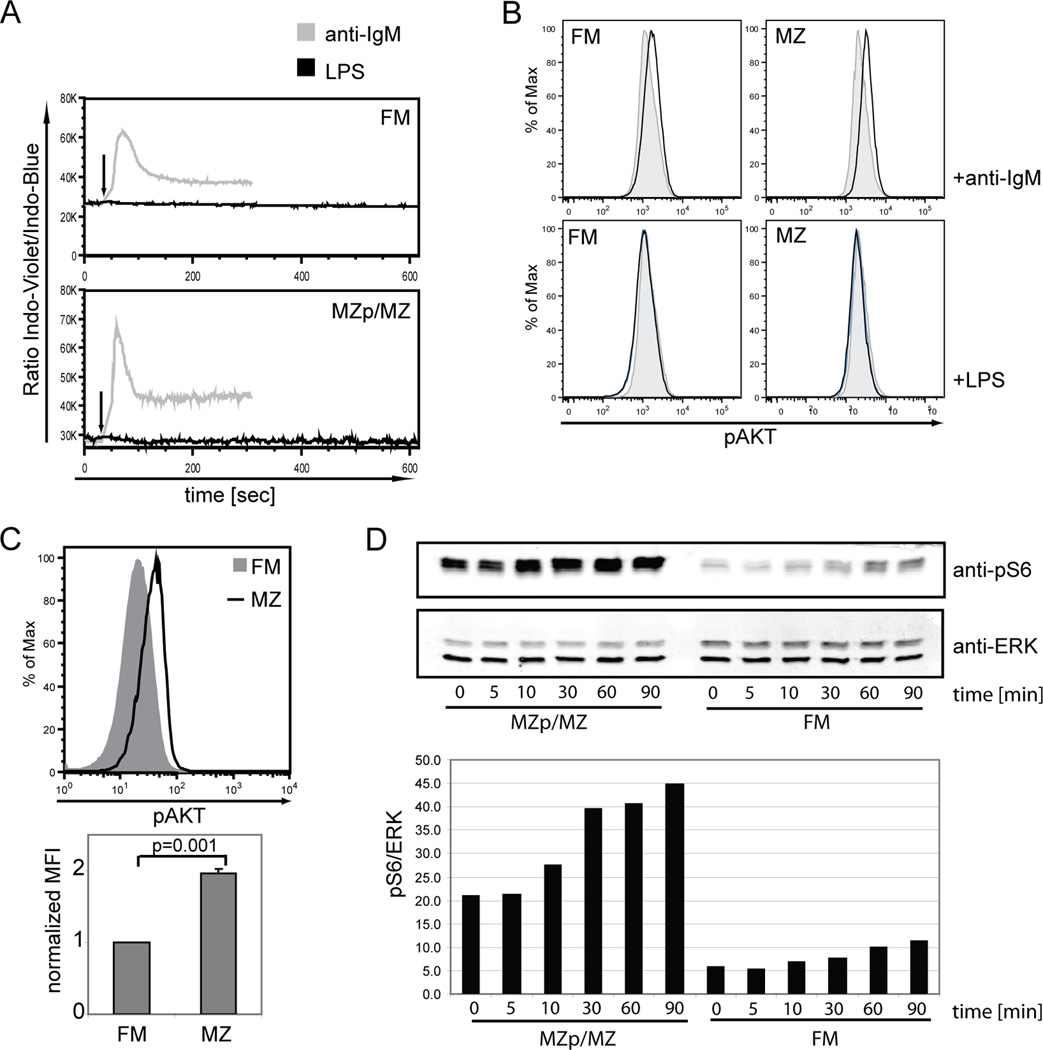
The splenic B cell compartment is comprised of two major, functionally distinct, mature B cell subsets, i.e., follicular mature (FM) and marginal zone (MZ) B cells. Whereas MZ B cells exhibit a robust proliferative response following stimulation with the TLR4 ligand LPS, FM B cells display markedly delayed and reduced levels of proliferation to the identical stimulus. The current study was designed to identify a potential mechanism(s) accounting for this differential responsiveness. In contrast to the delay in cell cycle entry, FM and MZ B cells exhibited nearly identical LPS-driven alterations in the expression level of cell surface activation markers. Furthermore, both the NF-kappaB and mTOR signaling cascades were similarly activated by LPS stimulation in FM vs MZ B cells, while inducible activation of ERK and AKT were nearly absent in both subsets. MZ B cells, however, exhibited higher basal levels of phospho-AKT and pS6, consistent with a preactivated status. Importantly, both basal and LPS activation-induced c-myc expression was markedly reduced in FM vs MZ B cells and enforced c-myc expression fully restored the defective proliferative response in FM B cells. These data support a model wherein TLR responses in FM B cells are tightly regulated by limiting c-myc levels, thereby providing an important checkpoint to control nonspecific FM B cell activation in the absence of cognate Ag.
Figures
References
-
- Martin F, Kearney JF. B-cell subsets and the mature preimmune repertoire. Marginal zone and B1 B cells as part of a "natural immune memory". Immunol Rev. 2000;175:70–79. - PubMed
-
- Saito T, Chiba S, Ichikawa M, Kunisato A, Asai T, Shimizu K, Yamaguchi T, Yamamoto G, Seo S, Kumano K, Nakagami-Yamaguchi E, Hamada Y, Aizawa S, Hirai H. Notch2 is preferentially expressed in mature B cells and indispensable for marginal zone B lineage development. Immunity. 2003;18:675–685. - PubMed
-
- Bendelac A, Bonneville M, Kearney JF. Autoreactivity by design: innate B and T lymphocytes. Nat Rev Immunol. 2001;1:177–186. - PubMed
-
- Oliver AM, Martin F, Gartland GL, Carter RH, Kearney JF. Marginal zone B cells exhibit unique activation, proliferative and immunoglobulin secretory responses. Eur J Immunol. 1997;27:2366–2374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
