

The lymphotoxin LTalpha(1)beta(2) controls postnatal and adult spleen marginal sinus vascular structure and function
- PMID: 19303389
- PMCID: PMC2874947
- DOI: 10.1016/j.immuni.2009.01.010
The lymphotoxin LTalpha(1)beta(2) controls postnatal and adult spleen marginal sinus vascular structure and function
Abstract
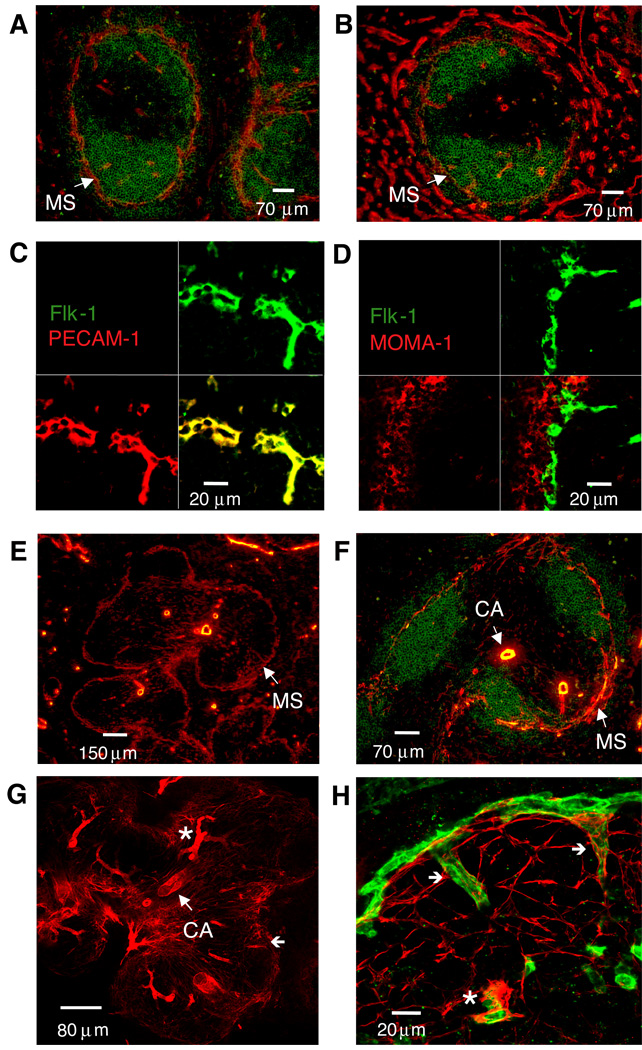
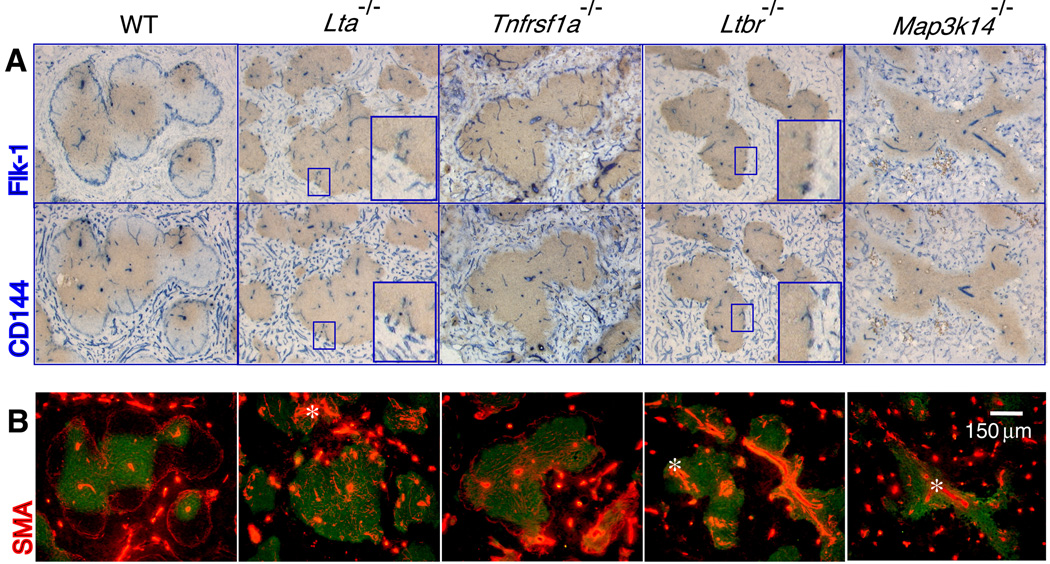
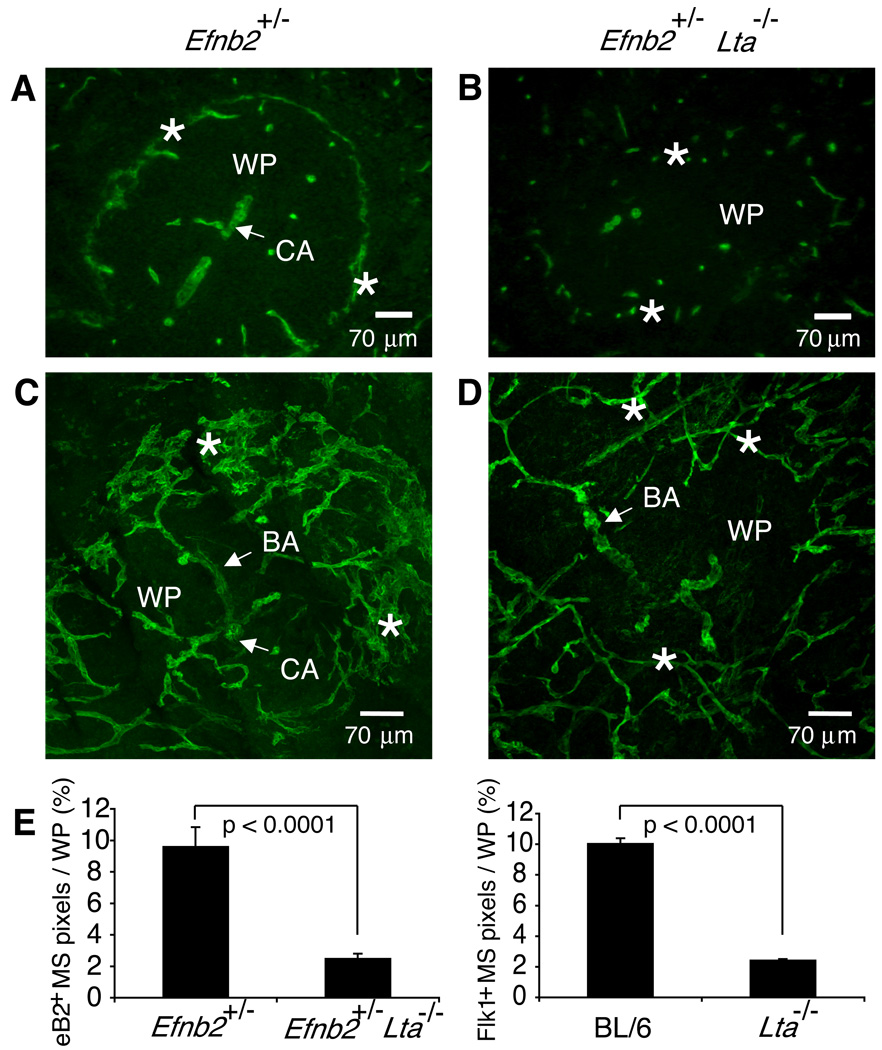
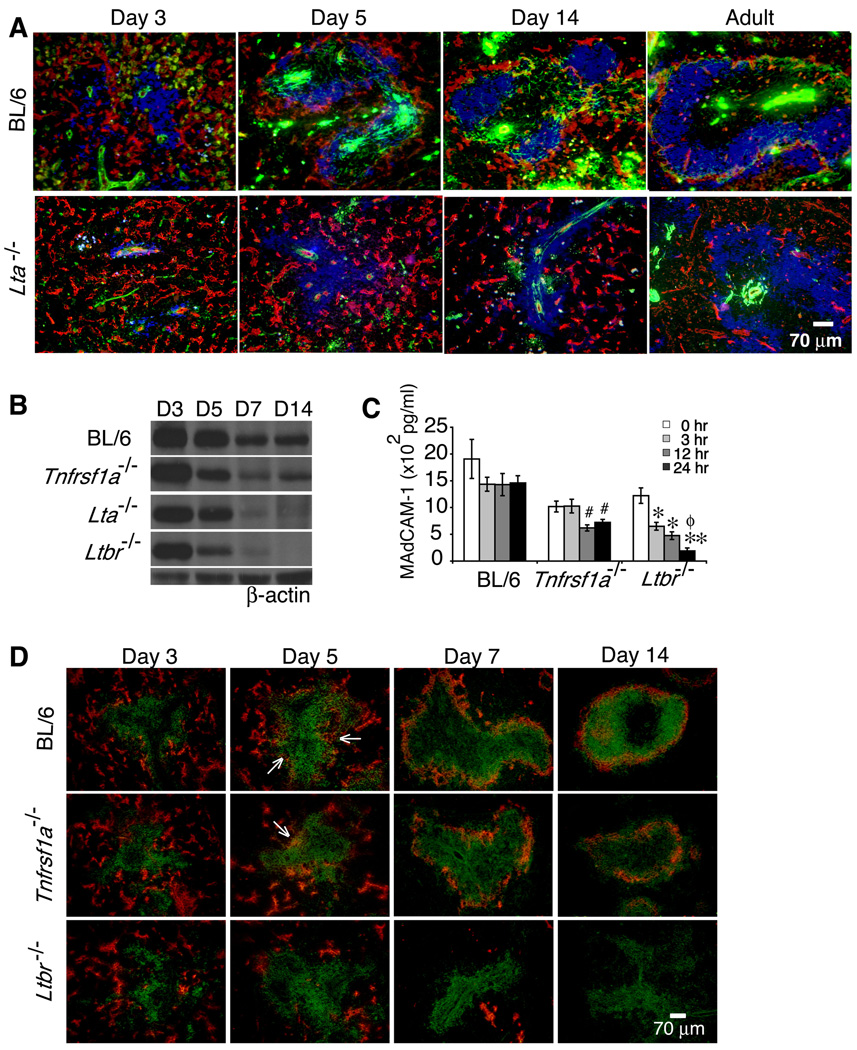
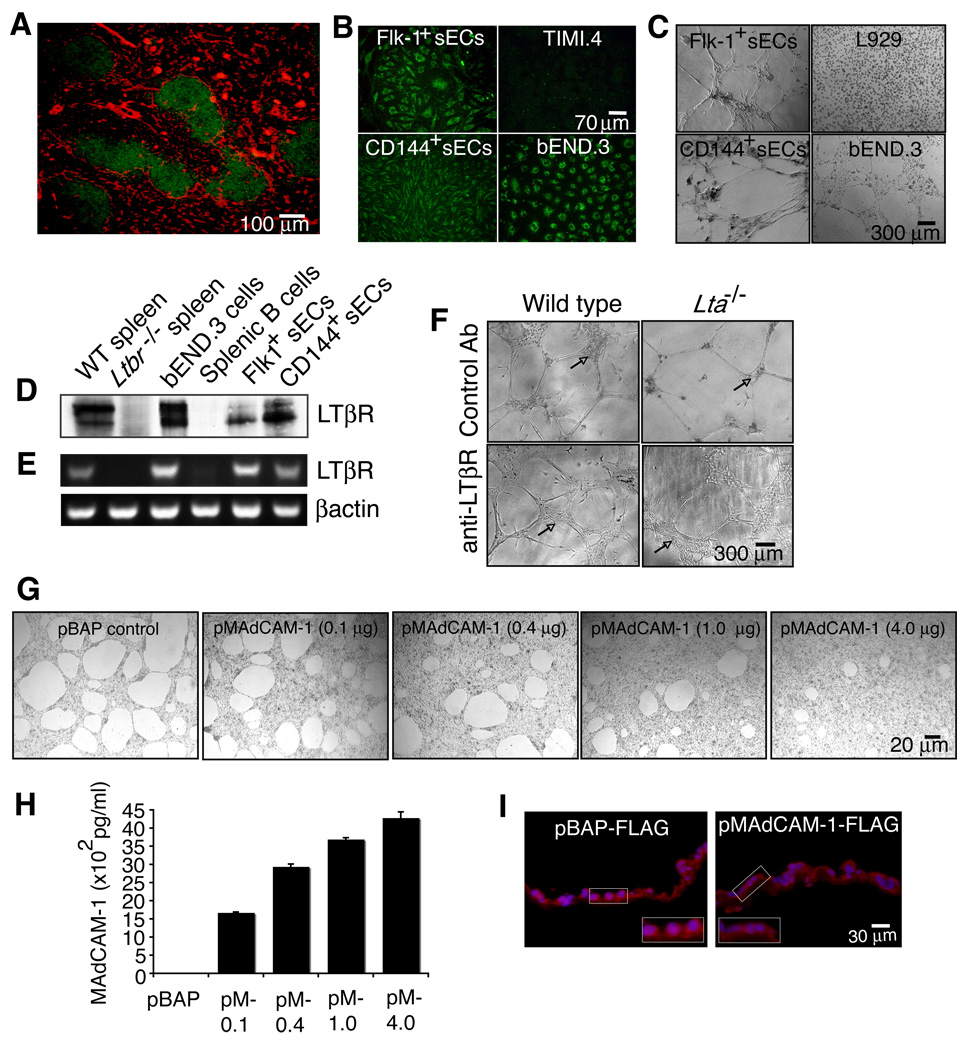
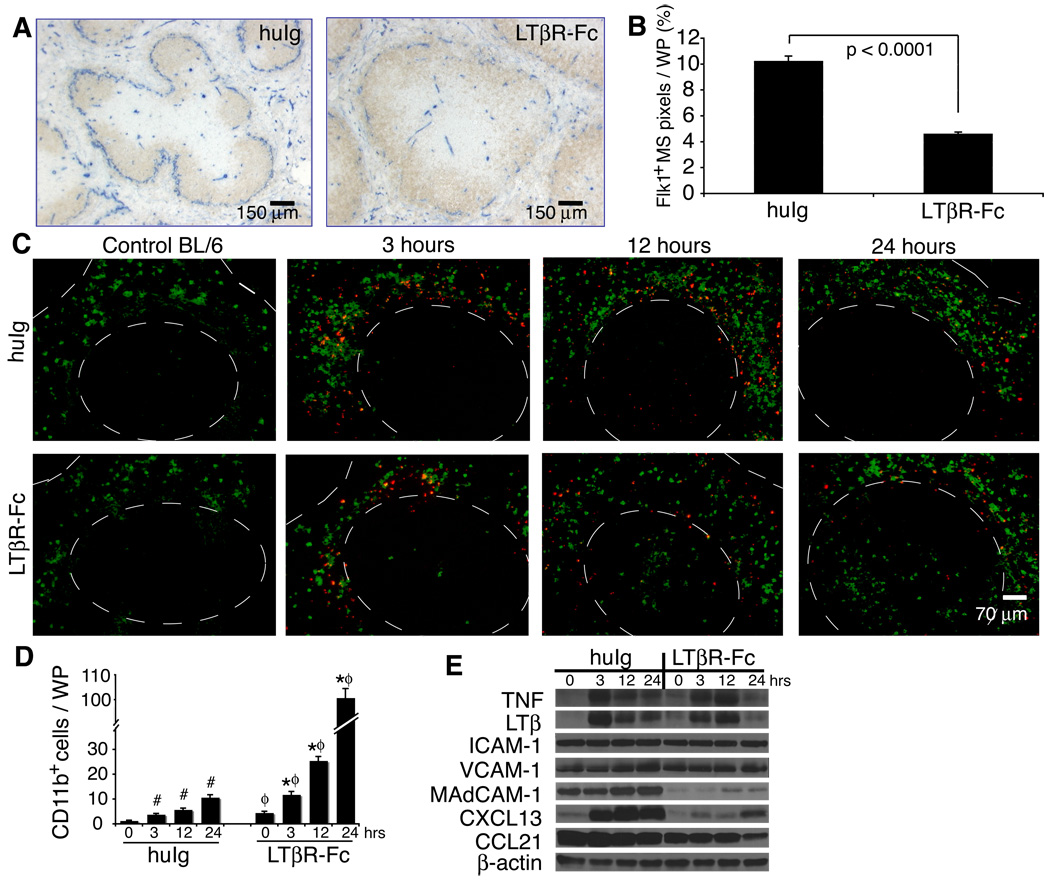
The lymphotoxin LTalpha(1)beta(2) supports the development and maintenance of several aspects of spleen structure, but its significance for marginal sinus (MS) vascular organization is unclear. We showed here that, in early postnatal lymphotoxin-deficient mice, the developing Flk-1+ white pulp vessels failed to organize or upregulate MAdCAM-1, leading to altered spatial rearrangement of both the white pulp endothelial cells and the smooth muscle actin-expressing cells. In vitro, MAdCAM-1 directed the reorganization of LTbeta receptor+ endothelial cells grown on Matrigel. LTalpha(1)beta(2) also regulated the maintenance of both MAdCAM-1 expression and mature MS structure in adult mice, contributing importantly to normal trafficking of CD11b+ cells in response to bacterial antigens. Together, our studies demonstrate that LTalpha(1)beta(2) and LTbeta receptor signals control proper development and maintenance of the mature MS structure and implicate MAdCAM-1 in the structuring of the MS endothelial cells that is important for the movement of immune cells within the spleen.
Figures
References
-
- Balazs M, Horvath G, Grama L, Balogh P. Phenotypic identification and development of distinct microvascular compartments in the postnatal mouse spleen. Cell Immunol. 2001;212:126–137. - PubMed
-
- Balazs M, Martin F, Zhou T, Kearney J. Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Immunity. 2002;17:341–352. - PubMed
-
- Balogh P, Balazs M, Czompoly T, Weih DS, Arnold HH, Weih F. Distinct roles of lymphotoxin-beta signaling and the homeodomain transcription factor Nkx2.3 in the ontogeny of endothelial compartments in spleen. Cell Tissue Res. 2007;328:473–486. - PubMed
-
- Banks TA, Rouse BT, Kerley MK, Blair PJ, Godfrey VL, Kuklin NA, Bouley DM, Thomas J, Kanangat S, Mucenski ML. Lymphotoxin-alpha-deficient mice. Effects on secondary lymphoid organ development and humoral immune responsiveness. J Immunol. 1995;155:1685–1693. - PubMed
-
- Bohnsack JF, Brown EJ. The role of the spleen in resistance to infection. Annu Rev Med. 1986;37:49–59. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
