

The BRC repeats of BRCA2 modulate the DNA-binding selectivity of RAD51
- PMID: 19303847
- PMCID: PMC2669112
- DOI: 10.1016/j.cell.2009.02.019
The BRC repeats of BRCA2 modulate the DNA-binding selectivity of RAD51
Abstract
The breast cancer susceptibility protein, BRCA2, is essential for recombinational DNA repair. BRCA2 delivers RAD51 to double-stranded DNA (dsDNA) breaks through interaction with eight conserved, approximately 35 amino acid motifs, the BRC repeats. Here we show that the solitary BRC4 promotes assembly of RAD51 onto single-stranded DNA (ssDNA), but not dsDNA, to stimulate DNA strand exchange. BRC4 acts by blocking ATP hydrolysis and thereby maintaining the active ATP-bound form of the RAD51-ssDNA filament. Single-molecule visualization shows that BRC4 does not disassemble RAD51-dsDNA filaments but rather blocks nucleation of RAD51 onto dsDNA. Furthermore, this behavior is manifested by a domain of BRCA2 comprising all eight BRC repeats. These results establish that the BRC repeats modulate RAD51-DNA interaction in two opposing but functionally reinforcing ways: targeting active RAD51 to ssDNA and prohibiting RAD51 nucleation onto dsDNA. Thus, BRCA2 recruits RAD51 to DNA breaks and, we propose, the BRC repeats regulate DNA-binding selectivity.
Figures

 ); ADP, Mg2+ + Ca2+ (
); ADP, Mg2+ + Ca2+ ( ), ATP and Ca2+ (
), ATP and Ca2+ ( ); AMP-PNP, Mg2+ + Ca2+ (
); AMP-PNP, Mg2+ + Ca2+ ( ); or no nucleotide cofactor, Mg2+ + Ca2+ (
); or no nucleotide cofactor, Mg2+ + Ca2+ ( ). Data obtained with the control mutant, GST-T1526A, ATP, Mg2+ + Ca2+: (
). Data obtained with the control mutant, GST-T1526A, ATP, Mg2+ + Ca2+: ( ). (C) BRC4 forms a stoichiometric complex with RAD51-ssDNA complexes. RAD51 (0.15 μM) was incubated with BRC4 () or BRCA2BRC1-8 (), or RAD51 (0.3 μM) was incubated with BRC4 (●), prior to incubation with 32P-labeled dT40 ssDNA (15 nM) for another 15 min in the presence of ATP, Mg2+ and Ca2+. Due to limitations of the protein stock, higher BRCA2BRC1-8 concentrations could not be examined. (D) BRC4 increases the stability of RAD51-ssDNA complexes to increasing concentrations salt: complexes were formed with RAD51 (0.9 μM) alone (upper panel) or RAD51 (0.9 μM) and GST-BRC4 (3.6 μM) (lower panel), in the presence of ADP and Mg2+.
). (C) BRC4 forms a stoichiometric complex with RAD51-ssDNA complexes. RAD51 (0.15 μM) was incubated with BRC4 () or BRCA2BRC1-8 (), or RAD51 (0.3 μM) was incubated with BRC4 (●), prior to incubation with 32P-labeled dT40 ssDNA (15 nM) for another 15 min in the presence of ATP, Mg2+ and Ca2+. Due to limitations of the protein stock, higher BRCA2BRC1-8 concentrations could not be examined. (D) BRC4 increases the stability of RAD51-ssDNA complexes to increasing concentrations salt: complexes were formed with RAD51 (0.9 μM) alone (upper panel) or RAD51 (0.9 μM) and GST-BRC4 (3.6 μM) (lower panel), in the presence of ADP and Mg2+.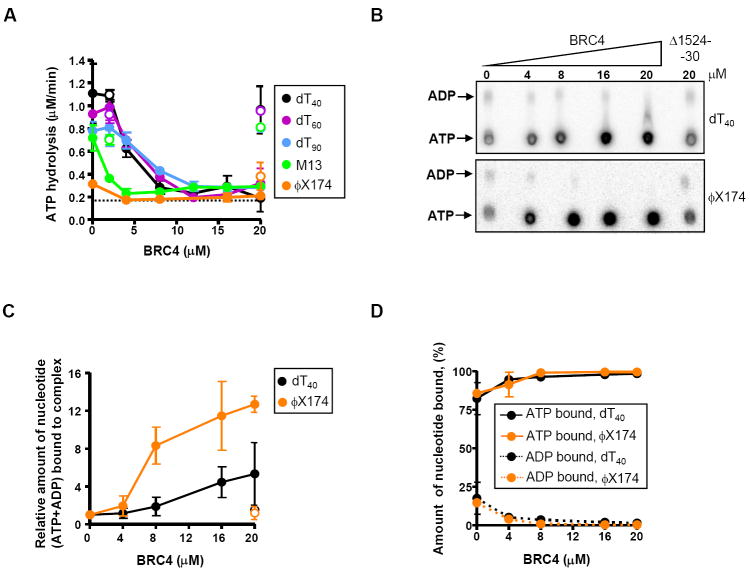
 ), dT90 (), M13 ssDNA (), or ϕX174 ssDNA (
), dT90 (), M13 ssDNA (), or ϕX174 ssDNA ( ), and further incubated for 1h in the presence of 0.5 mM ATP and 4 mM Mg2+. The dashed line represents the DNA-independent rate of ATP hydrolysis by RAD51 (~0.2 μM/min). (B) Nucleotides bound to purified nucleoprotein filaments as a function of BRC4 concentration as measured by TLC. (C) Amount of total nucleotide (ATP+ADP) bound to the RAD51-ssDNA complex formed with either dT40 (black circles) or ϕX174 ssDNA (orange circles) in presence of GST-BRC4 (filled circles) or GST-Δ1524-30 (open circles), relative to the amount in the absence of BRC4. (D) Percentages of ATP (solid lines) and ADP (dashed lines) in the RAD51 nucleoproteins filaments formed on dT40 (●) or ϕX174 ssDNA ().
), and further incubated for 1h in the presence of 0.5 mM ATP and 4 mM Mg2+. The dashed line represents the DNA-independent rate of ATP hydrolysis by RAD51 (~0.2 μM/min). (B) Nucleotides bound to purified nucleoprotein filaments as a function of BRC4 concentration as measured by TLC. (C) Amount of total nucleotide (ATP+ADP) bound to the RAD51-ssDNA complex formed with either dT40 (black circles) or ϕX174 ssDNA (orange circles) in presence of GST-BRC4 (filled circles) or GST-Δ1524-30 (open circles), relative to the amount in the absence of BRC4. (D) Percentages of ATP (solid lines) and ADP (dashed lines) in the RAD51 nucleoproteins filaments formed on dT40 (●) or ϕX174 ssDNA ().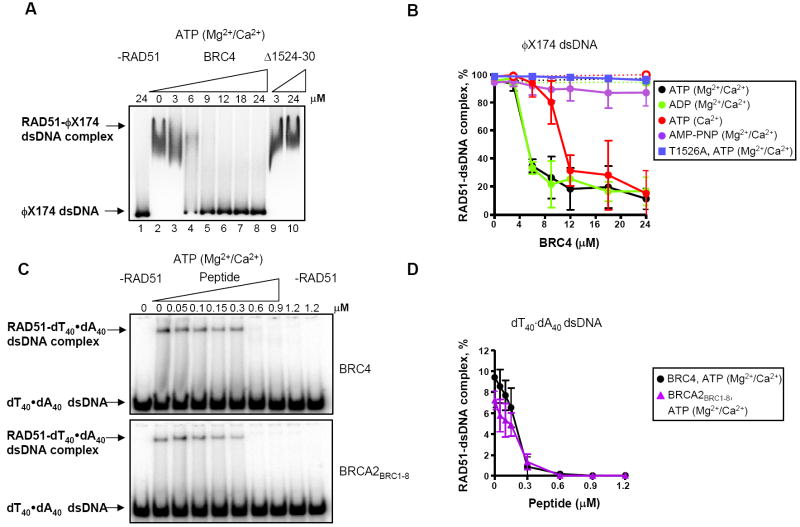 ); ATP and Ca2+ (); or AMP-PNP, Mg2+ + Ca2+ (). Filled circles and solid lines correspond to GST-BRC4; open circles and dashed lines correspond to GST-Δ1524-30. Data obtained with the control mutant, GST-T1526A, in the presence of ATP, Mg2+ + Ca2+ :(). (C) The effect of BRC4 or BRCA2BRC1-8 on RAD51-dsDNA complex formation using dA40·dT40 dsDNA. RAD51 (0.15 μM) was incubated with GST-BRC4 or BRCA2BRC1-8, at the concentrations indicated prior to incubation with 32P-labeled dA40·dT40 dsDNA (2.4 μM, nt) and further incubation for 1 h in the presence of ATP, Mg2+ and Ca2+. Protein-DNA complexes were resolved in 6% PAGE and analyzed by autoradiography. (D) Data obtained as in (C), quantified and plotted: GST-BRC4, (●); BRCA2BRC1-8 (
); ATP and Ca2+ (); or AMP-PNP, Mg2+ + Ca2+ (). Filled circles and solid lines correspond to GST-BRC4; open circles and dashed lines correspond to GST-Δ1524-30. Data obtained with the control mutant, GST-T1526A, in the presence of ATP, Mg2+ + Ca2+ :(). (C) The effect of BRC4 or BRCA2BRC1-8 on RAD51-dsDNA complex formation using dA40·dT40 dsDNA. RAD51 (0.15 μM) was incubated with GST-BRC4 or BRCA2BRC1-8, at the concentrations indicated prior to incubation with 32P-labeled dA40·dT40 dsDNA (2.4 μM, nt) and further incubation for 1 h in the presence of ATP, Mg2+ and Ca2+. Protein-DNA complexes were resolved in 6% PAGE and analyzed by autoradiography. (D) Data obtained as in (C), quantified and plotted: GST-BRC4, (●); BRCA2BRC1-8 ( ).
). ), 0.5 μM (), 250 nM (
), 0.5 μM (), 250 nM ( ), and 125 nM (
), and 125 nM ( ) GST-BRC4; or 0.5 μM (
) GST-BRC4; or 0.5 μM ( ) and 125 nM (
) and 125 nM ( ) BRCA2BRC1-8. The dashed lines represent the linear regressions of the data. (E) Rate of cluster formation vs. BRC4 or BRCA2BRC1-8 concentration; each time point is the average of 4–9 individual molecules analyzed as in D.
) BRCA2BRC1-8. The dashed lines represent the linear regressions of the data. (E) Rate of cluster formation vs. BRC4 or BRCA2BRC1-8 concentration; each time point is the average of 4–9 individual molecules analyzed as in D.


References
-
- Baumann P, Benson FE, West SC. Human Rad51 protein promotes ATP-dependent homologous pairing and strand transfer reactions in vitro. Cell. 1996;87:757–766. - PubMed
-
- Benson FE, Baumann P, West SC. Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature. 1998;391:401–404. - PubMed
-
- Bianco PR, Brewer LR, Corzett M, Balhorn R, Yeh Y, Kowalczykowski SC, Baskin RJ. Processive translocation and DNA unwinding by individual RecBCD enzyme molecules. Nature. 2001;409:374–378. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
