

A phosphoinositide switch controls the maturation and signaling properties of APPL endosomes
- PMID: 19303853
- PMCID: PMC2705806
- DOI: 10.1016/j.cell.2009.01.032
A phosphoinositide switch controls the maturation and signaling properties of APPL endosomes
Abstract
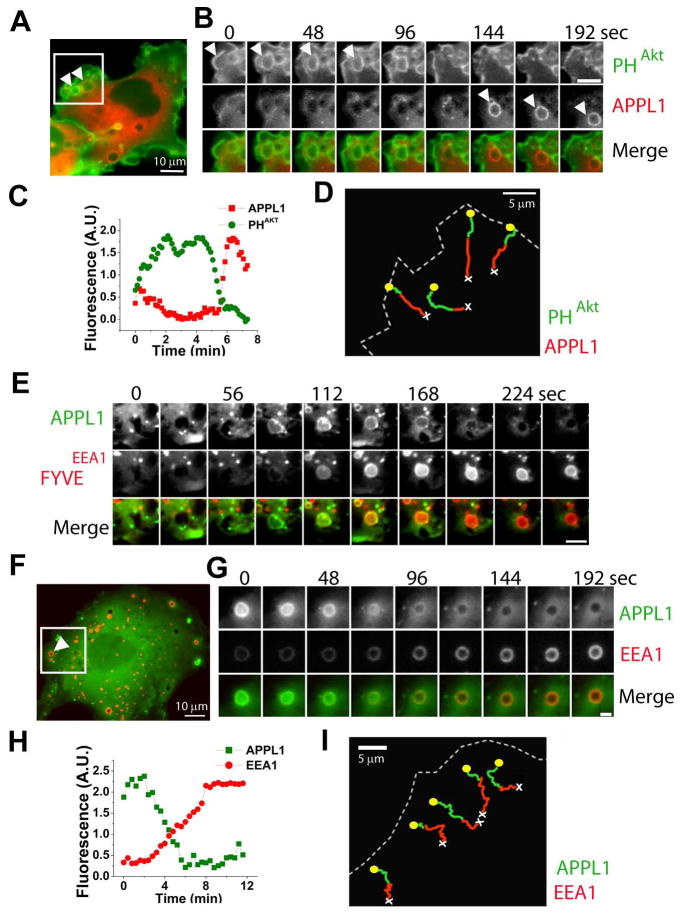
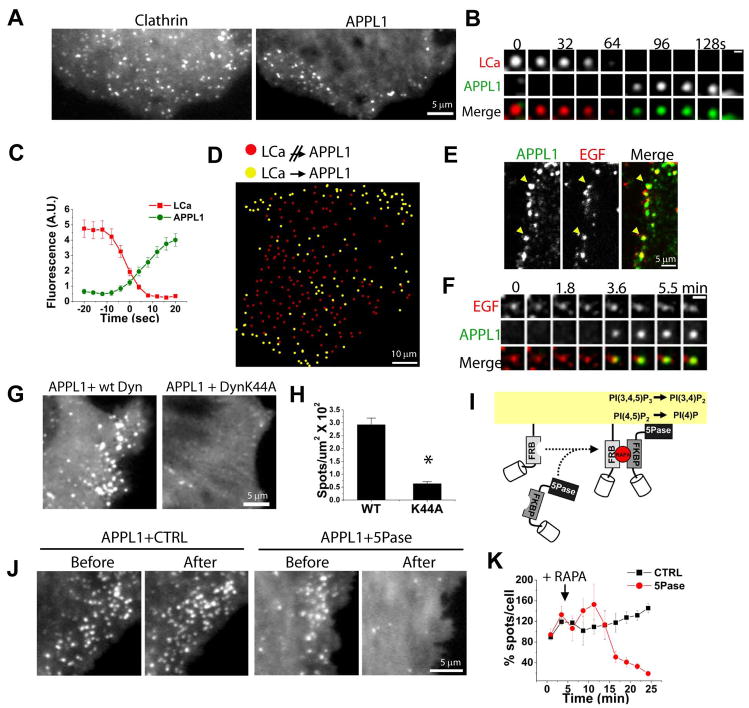
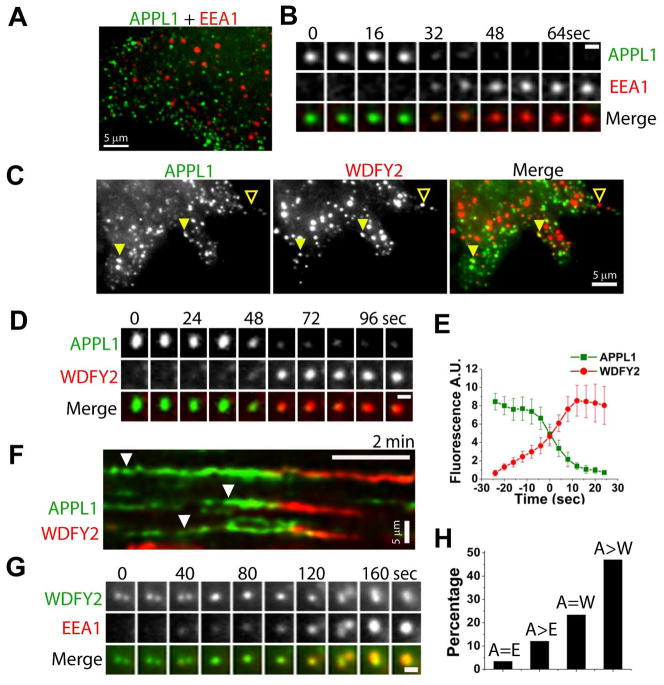
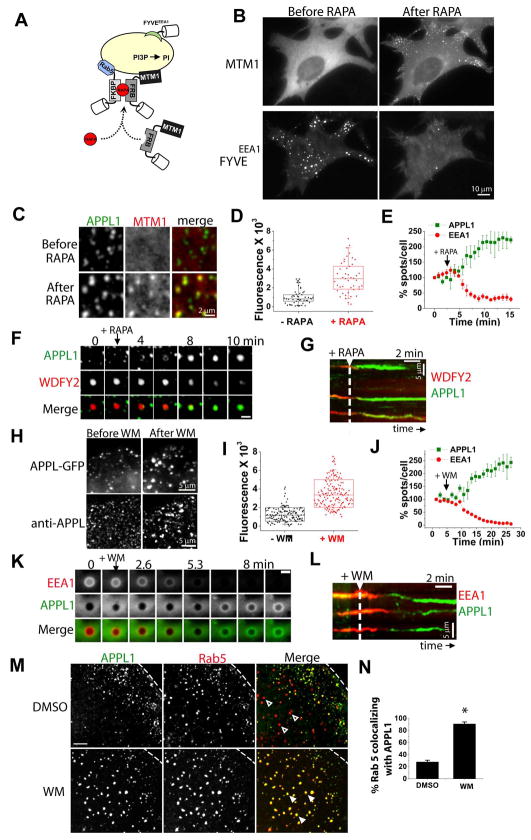
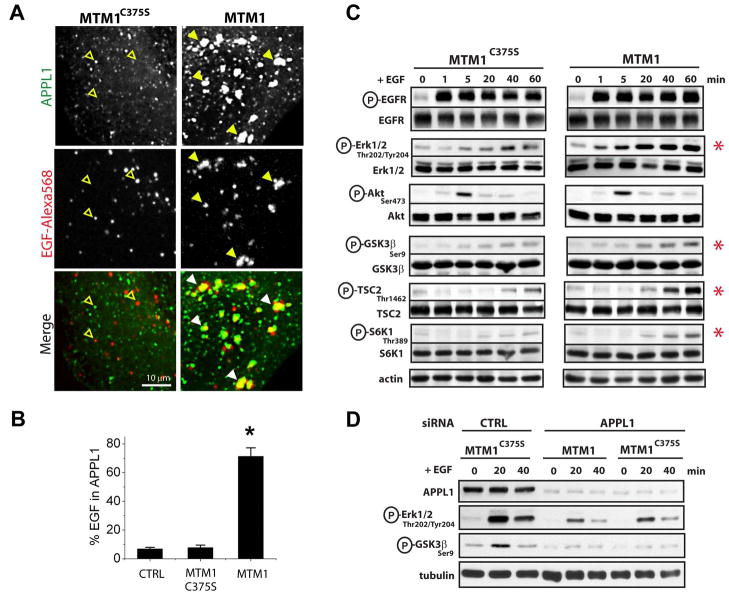
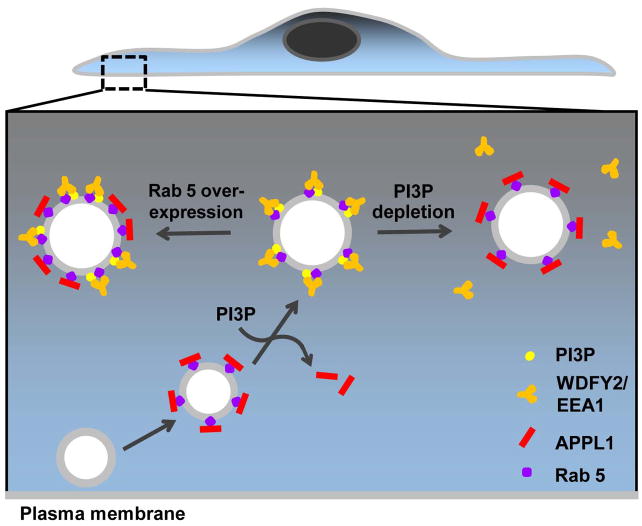
The recent identification of several novel endocytic compartments has challenged our current understanding of the topological and functional organization of the endocytic pathway. Using quantitative single vesicle imaging and acute manipulation of phosphoinositides we show that APPL endosomes, which participate in growth factor receptor trafficking and signaling, represent an early endocytic intermediate common to a subset of clathrin derived endocytic vesicles and macropinosomes. Most APPL endosomes are precursors of classical PI3P positive endosomes, and PI3P plays a critical role in promoting this conversion. Depletion of PI3P causes a striking reversion of Rab5 positive endosomes to the APPL stage, and results in enhanced growth factor signaling. These findings reveal a surprising plasticity of the early endocytic pathway. Importantly, PI3P functions as a switch to dynamically regulate maturation and signaling of APPL endosomes.
Figures
References
-
- Behnia R, Munro S. Organelle identity and the signposts for membrane traffic. Nature. 2005;438:597–604. - PubMed
-
- Colaluca IN, Tosoni D, Nuciforo P, Senic-Matuglia F, Galimberti V, Viale G, Pece S, Di Fiore PP. NUMB controls p53 tumour suppressor activity. Nature. 2008;451:76–80. - PubMed
-
- De Matteis MA, Di Campli A, Godi A. The role of the phosphoinositides at the Golgi complex. Biochim Biophys Acta. 2005;1744:396–405. - PubMed
-
- Di Fiore PP, De Camilli P. Endocytosis and signaling. an inseparable partnership. Cell. 2001;106:1–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 5T32GM07205/GM/NIGMS NIH HHS/United States
- P01 CA046128/CA/NCI NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P30 DA018343/DA/NIDA NIH HHS/United States
- DK45735/DK/NIDDK NIH HHS/United States
- CA46128/CA/NCI NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- DA018343/DA/NIDA NIH HHS/United States
- DP2 OD002980/OD/NIH HHS/United States
- 1DP20D002980-01/DP/NCCDPHP CDC HHS/United States
- NS36251/NS/NINDS NIH HHS/United States
- R37 NS036251/NS/NINDS NIH HHS/United States
- R01 NS036251/NS/NINDS NIH HHS/United States
- T32 GM007205/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
