

G clustering is important for the initiation of transcription-induced R-loops in vitro, whereas high G density without clustering is sufficient thereafter
- PMID: 19307304
- PMCID: PMC2682002
- DOI: 10.1128/MCB.00139-09
G clustering is important for the initiation of transcription-induced R-loops in vitro, whereas high G density without clustering is sufficient thereafter
Abstract
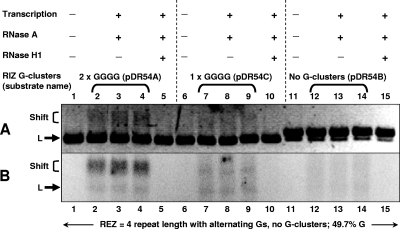
R-loops form cotranscriptionally in vitro and in vivo at transcribed duplex DNA regions when the nascent RNA is G-rich, particularly with G clusters. This is the case for phage polymerases, as used here (T7 RNA polymerase), as well as RNA polymerases in bacteria, Saccharomyces cerevisiae, avians, mice, and humans. The nontemplate strand is left in a single-stranded configuration within the R-loop region. These structures are known to form at mammalian immunoglobulin class switch regions, thus exposing regions of single-stranded DNA for the action of AID, a single-strand-specific cytidine deaminase. R-loops form by thread-back of the RNA onto the template DNA strand, and here we report that G clusters are extremely important for the initiation phase of R-loop formation. Even very short regions with one GGGG sequence can initiate R-loops much more efficiently than random sequences. The high efficiencies observed with G clusters cannot be achieved by having a very high G density alone. Annealing of the transcript, which is otherwise disadvantaged relative to the nontemplate DNA strand because of unfavorable proximity while exiting the RNA polymerase, can offer greater stability if it occurs at the G clusters, thereby initiating an R-loop. R-loop elongation beyond the initiation zone occurs in a manner that is not as reliant on G clusters as it is on a high G density. These results lead to a model in which G clusters are important to nucleate the thread-back of RNA for R-loop initiation and, once initiated, the elongation of R-loops is primarily determined by the density of G on the nontemplate DNA strand. Without both a favorable R-loop initiation zone and elongation zone, R-loop formation is inefficient.
Figures






Similar articles
-
Competition between the RNA transcript and the nontemplate DNA strand during R-loop formation in vitro: a nick can serve as a strong R-loop initiation site.Mol Cell Biol. 2010 Jan;30(1):146-59. doi: 10.1128/MCB.00897-09. Mol Cell Biol. 2010. PMID: 19841062 Free PMC article.
-
Detection and Characterization of R Loop Structures.Methods Mol Biol. 2017;1543:231-242. doi: 10.1007/978-1-4939-6716-2_13. Methods Mol Biol. 2017. PMID: 28349431
-
Intracellular transcription of G-rich DNAs induces formation of G-loops, novel structures containing G4 DNA.Genes Dev. 2004 Jul 1;18(13):1618-29. doi: 10.1101/gad.1200804. Genes Dev. 2004. PMID: 15231739 Free PMC article.
-
New insight into the biology of R-loops.Mutat Res. 2020 May-Dec;821:111711. doi: 10.1016/j.mrfmmm.2020.111711. Epub 2020 May 28. Mutat Res. 2020. PMID: 32516653 Review.
-
R-loop structure: the formation and the effects on genomic stability.Yi Chuan. 2014 Dec;36(12):1185-94. doi: 10.3724/SP.J.1005.2014.1185. Yi Chuan. 2014. PMID: 25487262 Review.
Cited by
-
R-loop: an emerging regulator of chromatin dynamics.Acta Biochim Biophys Sin (Shanghai). 2016 Jul;48(7):623-31. doi: 10.1093/abbs/gmw052. Epub 2016 Jun 1. Acta Biochim Biophys Sin (Shanghai). 2016. PMID: 27252122 Free PMC article. Review.
-
Capsid amino acids at positions 247 and 270 are involved in the virulence of betanodaviruses to European sea bass.Sci Rep. 2019 Oct 1;9(1):14068. doi: 10.1038/s41598-019-50622-1. Sci Rep. 2019. PMID: 31575937 Free PMC article.
-
Understanding Long Noncoding RNA and Chromatin Interactions: What We Know So Far.Noncoding RNA. 2019 Dec 3;5(4):54. doi: 10.3390/ncrna5040054. Noncoding RNA. 2019. PMID: 31817041 Free PMC article. Review.
-
Structure and Dynamics of DNA and RNA Double Helices Formed by d(CTG), d(GTC), r(CUG), and r(GUC) Trinucleotide Repeats and Associated DNA-RNA Hybrids.J Phys Chem B. 2023 Sep 21;127(37):7907-7924. doi: 10.1021/acs.jpcb.3c03538. Epub 2023 Sep 8. J Phys Chem B. 2023. PMID: 37681731 Free PMC article.
-
Characterization of R-Loop-Interacting Proteins in Embryonic Stem Cells Reveals Roles in rRNA Processing and Gene Expression.Mol Cell Proteomics. 2021;20:100142. doi: 10.1016/j.mcpro.2021.100142. Epub 2021 Aug 31. Mol Cell Proteomics. 2021. PMID: 34478875 Free PMC article.
References
-
- Aguilera, A., and B. Gomez-Gonzalez. 2008. Genome instability: a mechanistic view of its causes and consequences. Nat. Rev. Genet. 9204-217. - PubMed
-
- Artsimovitch, I., and R. Landick. 2002. The transcriptional regulator RfaH stimulates RNA chain synthesis after recruitment to elongation complexes by the exposed nontemplate DNA strand. Cell 109193-203. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources