

The Wiskott-Aldrich syndrome protein is required for iNKT cell maturation and function
- PMID: 19307326
- PMCID: PMC2715111
- DOI: 10.1084/jem.20081773
The Wiskott-Aldrich syndrome protein is required for iNKT cell maturation and function
Abstract
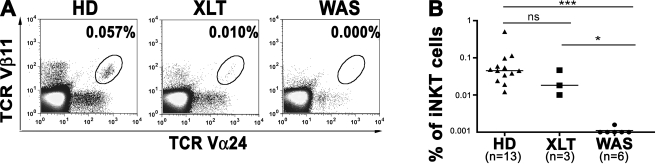
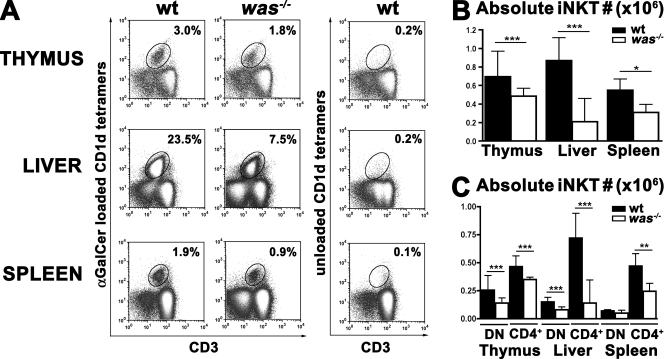
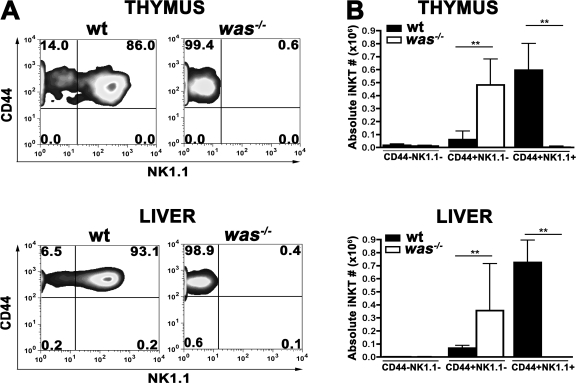
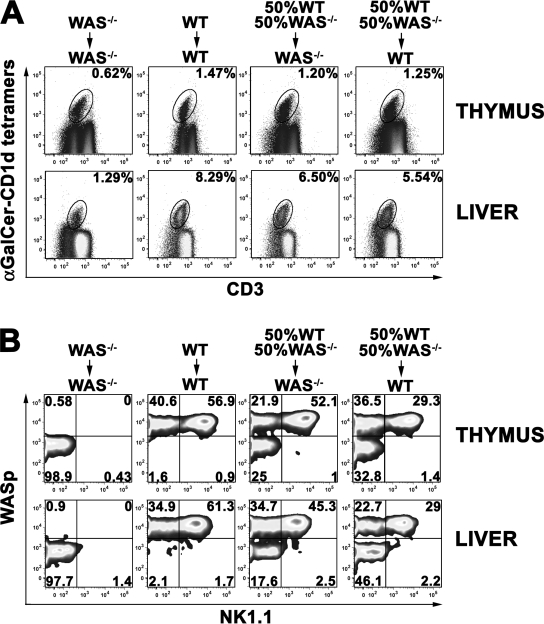
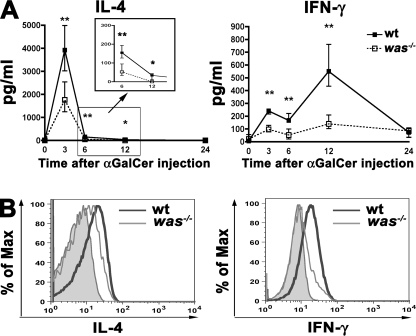
The Wiskott-Aldrich syndrome (WAS) protein (WASp) is a regulator of actin cytoskeleton in hematopoietic cells. Mutations of the WASp gene cause WAS. Although WASp is involved in various immune cell functions, its role in invariant natural killer T (iNKT) cells has never been investigated. Defects of iNKT cells could indeed contribute to several WAS features, such as recurrent infections and high tumor incidence. We found a profound reduction of circulating iNKT cells in WAS patients, directly correlating with the severity of clinical phenotype. To better characterize iNKT cell defect in the absence of WASp, we analyzed was(-/-) mice. iNKT cell numbers were significantly reduced in the thymus and periphery of was(-/-) mice as compared with wild-type controls. Moreover analysis of was(-/-) iNKT cell maturation revealed a complete arrest at the CD44(+) NK1.1(-) intermediate stage. Notably, generation of BM chimeras demonstrated a was(-/-) iNKT cell-autonomous developmental defect. was(-/-) iNKT cells were also functionally impaired, as suggested by the reduced secretion of interleukin 4 and interferon gamma upon in vivo activation. Altogether, these results demonstrate the relevance of WASp in integrating signals critical for development and functional differentiation of iNKT cells and suggest that defects in these cells may play a role in WAS pathology.
Figures
References
-
- Bendelac A., Savage P.B., Teyton L. 2007. The biology of NKT cells.Annu. Rev. Immunol. 25:297–336 - PubMed
-
- Godfrey D.I., Berzins S.P. 2007. Control points in NKT-cell development.Nat. Rev. Immunol. 7:505–518 - PubMed
-
- Tupin E., Kinjo Y., Kronenberg M. 2007. The unique role of natural killer T cells in the response to microorganisms.Nat. Rev. Microbiol. 5:405–417 - PubMed
-
- Van Kaer L. 2007. NKT cells: T lymphocytes with innate effector functions.Curr. Opin. Immunol. 19:354–364 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
