

A role for human skin-resident T cells in wound healing
- PMID: 19307328
- PMCID: PMC2715110
- DOI: 10.1084/jem.20081787
A role for human skin-resident T cells in wound healing
Abstract
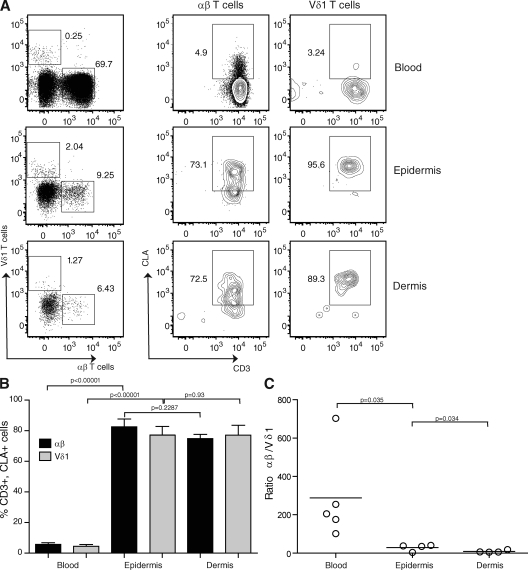
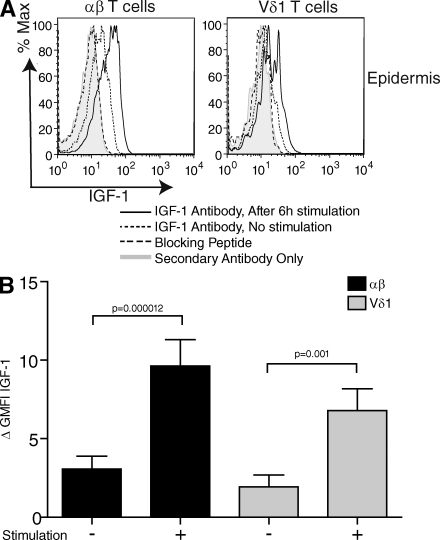
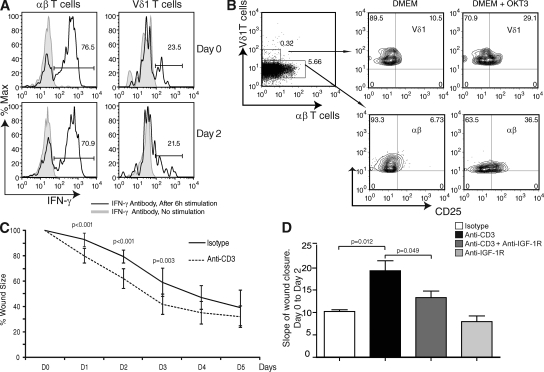
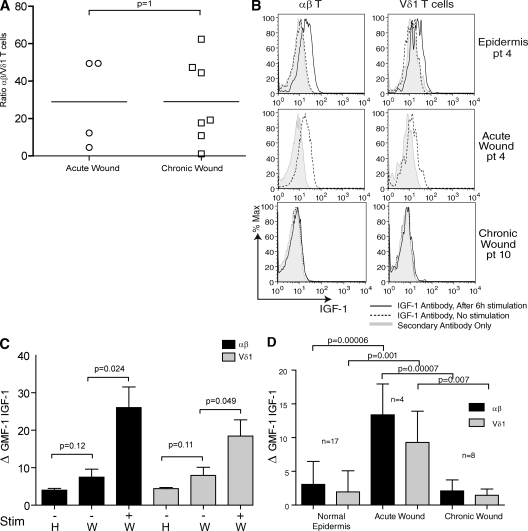
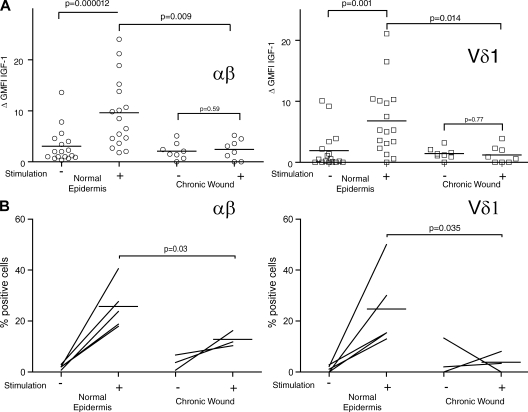
Epidermal T cells have been shown to play unique roles in tissue homeostasis and repair in mice through local secretion of distinct growth factors in the skin. Human epidermis contains both alphabeta(+) and gammadelta(+) T cells whose functional capabilities are not understood. We demonstrate that human epidermal T cells are able to produce insulin-like growth factor 1 (IGF-1) upon activation and promote wound healing in a skin organ culture model. Moreover, an analysis of the functional capabilities of T cells isolated from acute versus chronic wounds revealed a striking difference. Both alphabeta(+) and Vdelta1(+) T cells isolated from acute wounds actively produced IGF-1, demonstrating that they are activated during tissue damage to participate in wound repair. In contrast, IGF-1 production could not be detected in T cells isolated from chronic wounds. In fact, skin T cells isolated from chronic wounds were refractory to further stimulation, suggesting an unresponsive state. Collectively, these results define a novel role for human epidermis-resident T cells in wound healing and provide new insight into our understanding of chronic wound persistence.
Figures
References
-
- Kuehn B.M. 2007. Chronic wound care guidelines issued.JAMA. 297:938–939 - PubMed
-
- Dupuy P., Heslan M., Fraitag S., Hercend T., Dubertret L., Bagot M. 1990. T-cell receptor-γδ bearing lymphocytes in normal and inflammatory human skin.J. Invest. Dermatol. 94:764–768 - PubMed
-
- Ebert L.M., Meuter S., Moser B. 2006. Homing and function of human skin γδ T cells and NK cells: relevance for tumor surveillance.J. Immunol. 176:4331–4336 - PubMed
-
- Clark R.A., Chong B., Mirchandani N., Brinster N.K., Yamanaka K., Dowgiert R.K., Kupper T.S. 2006. The vast majority of CLA+ T cells are resident in normal skin.J. Immunol. 176:4431–4439 - PubMed
-
- Tamaki K., Sugaya M., Tada Y., Yasaka N., Uehira M., Nishimoto H., Nakamura K. 2001. Epidermal and dermal γδ T cells.Chem. Immunol. 79:43–51 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
