

Activin receptor signaling regulates prostatic epithelial cell adhesion and viability
- PMID: 19308291
- PMCID: PMC2657882
- DOI: 10.1593/neo.81544
Activin receptor signaling regulates prostatic epithelial cell adhesion and viability
Abstract
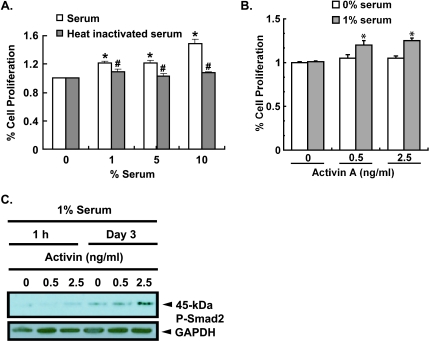
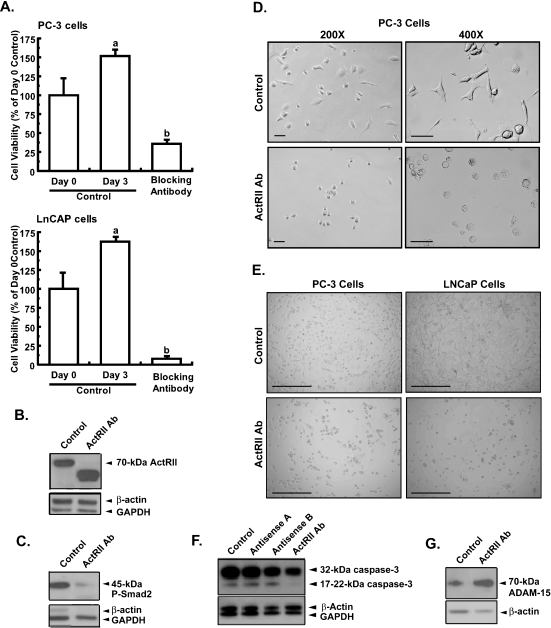
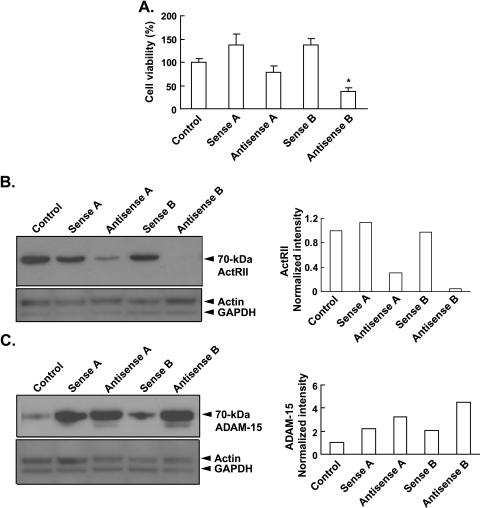
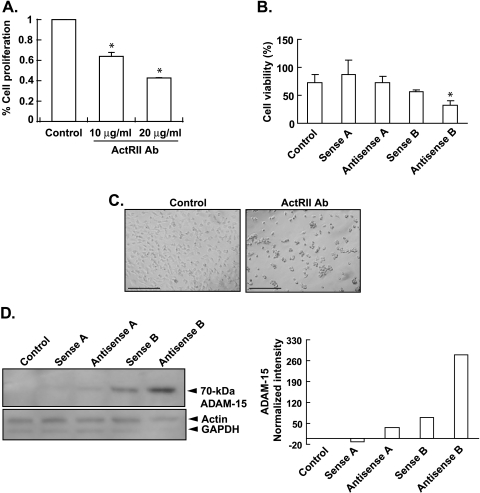
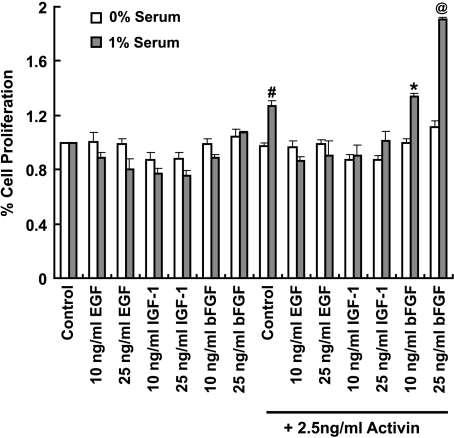
Mutational changes coupled with endocrine, paracrine, and/or autocrine signals regulate cell division during carcinogenesis. The hormone signals remain undefined, although the absolute requirement in vitro for fetal serum indicates the necessity for a fetal serum factor(s) in cell proliferation. Using prostatic cancer cell (PCC) lines as a model of cancer cell proliferation, we have identified the fetal serum component activin A and its signaling through the activin receptor type II (ActRII), as necessary, although not sufficient, for PCC proliferation. Activin A induced Smad2 phosphorylation and PCC proliferation, but only in the presence of fetal bovine serum (FBS). Conversely, activin A antibodies and inhibin A suppressed FBS-induced PCC proliferation confirming activin A as one of multiple serum components required for PCC proliferation. Basic fibroblast growth factor was subsequently shown to synergize activin A-induced PCC proliferation. Inhibition of ActRII signaling using a blocking antibody or antisense-P decreased mature ActRII expression, Smad2 phosphorylation, and the apparent viability of PCCs and neuroblastoma cells grown in FBS. Suppression of ActRII signaling in PCC and neuroblastoma cells did not induce apoptosis as indicated by the ratio of active/inactive caspase 3 but did correlate with increased cell detachment and ADAM-15 expression, a disintegrin whose expression is strongly correlated with prostatic metastasis. These findings indicate that ActRII signaling is required for PCC and neuroblastoma cell viability, with ActRII mediating cell fate via the regulation of cell adhesion. That ActRII signaling governs both cell viability and cell adhesion has important implications for developing therapeutic strategies to regulate cancer growth and metastasis.
Figures
Similar articles
-
An activin mutant with disrupted ALK4 binding blocks signaling via type II receptors.J Biol Chem. 2004 Jul 2;279(27):28036-44. doi: 10.1074/jbc.M402782200. Epub 2004 Apr 27. J Biol Chem. 2004. PMID: 15123686
-
Activin type II receptor signaling in cardiac aging and heart failure.Sci Transl Med. 2019 Mar 6;11(482):eaau8680. doi: 10.1126/scitranslmed.aau8680. Sci Transl Med. 2019. PMID: 30842316 Free PMC article.
-
Cripto forms a complex with activin and type II activin receptors and can block activin signaling.Proc Natl Acad Sci U S A. 2003 Apr 29;100(9):5193-8. doi: 10.1073/pnas.0531290100. Epub 2003 Apr 7. Proc Natl Acad Sci U S A. 2003. PMID: 12682303 Free PMC article.
-
Activin signaling and its role in regulation of cell proliferation, apoptosis, and carcinogenesis.Exp Biol Med (Maywood). 2006 May;231(5):534-44. doi: 10.1177/153537020623100507. Exp Biol Med (Maywood). 2006. PMID: 16636301 Review.
-
Activins, inhibins, and follistatins: from endocrinology to signaling. A paradigm for the new millennium.Exp Biol Med (Maywood). 2002 Oct;227(9):724-52. doi: 10.1177/153537020222700905. Exp Biol Med (Maywood). 2002. PMID: 12324653 Review.
Cited by
-
Identification of a bone morphogenetic protein type 2 receptor neutralizing antibody.BMC Res Notes. 2019 Jun 11;12(1):331. doi: 10.1186/s13104-019-4367-0. BMC Res Notes. 2019. PMID: 31186065 Free PMC article.
-
The War on Cancer rages on.Neoplasia. 2009 Dec;11(12):1252-63. doi: 10.1593/neo.91866. Neoplasia. 2009. PMID: 20019833 Free PMC article.
-
SETD7 functions as a transcription repressor in prostate cancer via methylating FOXA1.Proc Natl Acad Sci U S A. 2023 Aug 15;120(33):e2220472120. doi: 10.1073/pnas.2220472120. Epub 2023 Aug 7. Proc Natl Acad Sci U S A. 2023. PMID: 37549269 Free PMC article.
-
Expression of nodal and nodal receptors in prostate stem cells and prostate cancer cells: autocrine effects on cell proliferation and migration.Prostate. 2011 Jul;71(10):1084-96. doi: 10.1002/pros.21326. Epub 2011 Jan 12. Prostate. 2011. PMID: 21557273 Free PMC article.
-
Serum follistatin in patients with prostate cancer metastatic to the bone.Clin Exp Metastasis. 2010 Dec;27(8):549-55. doi: 10.1007/s10585-010-9344-x. Epub 2010 Jul 11. Clin Exp Metastasis. 2010. PMID: 20623366
References
-
- McCullough KD, Coleman WB, Smith GJ, Grisham JW. Age-dependent induction of hepatic tumor regression by the tissue microenvironment after transplantation of neoplastically transformed rat liver epithelial cells into the liver. Cancer Res. 1997;57:1807–1813. - PubMed
-
- Coleman WB, McCullough KD, Esch GL, Faris RA, Hixson DC, Smith GJ, Grisham JW. Evaluation of the differentiation potential of WB-F344 rat liver epithelial stem-like cells in vivo. Differentiation to hepatocytes after transplantation into dipeptidylpeptidase-IV-deficient rat liver. Am J Pathol. 1997;151:353–359. - PMC - PubMed
-
- Lee LM, Seftor EA, Bonde G, Cornell RA, Hendrix MJ. The fate of human malignant melanoma cells transplanted into zebrafish embryos: assessment of migration and cell division in the absence of tumor formation. Dev Dyn. 2005;233:1560–1570. - PubMed
-
- Topczewska JM, Postovit LM, Margaryan NV, Sam A, Hess AR, Wheaton WW, Nickoloff BJ, Topczewski J, Hendrix MJ. Embryonic and tumorigenic pathways converge via Nodal signaling: role in melanoma aggressiveness. Nat Med. 2006;12:925–932. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials