

Chronic vagus nerve stimulation induces neuronal plasticity in the rat hippocampus
- PMID: 19309534
- PMCID: PMC2879889
- DOI: 10.1017/S1461145709000200
Chronic vagus nerve stimulation induces neuronal plasticity in the rat hippocampus
Abstract
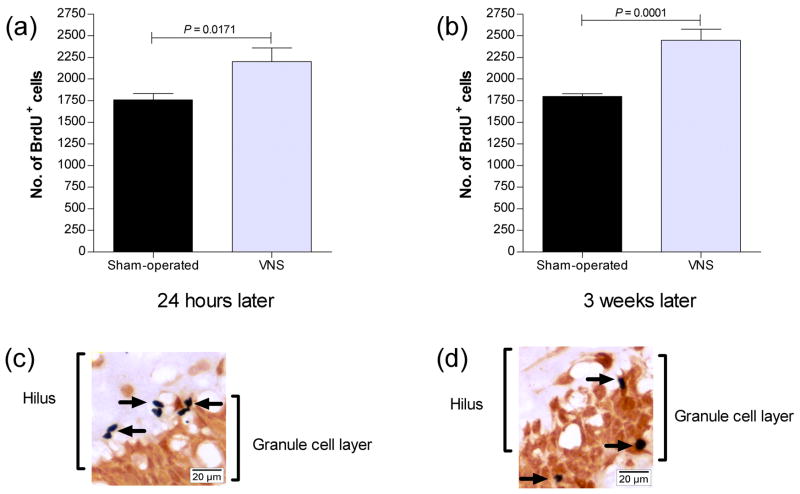
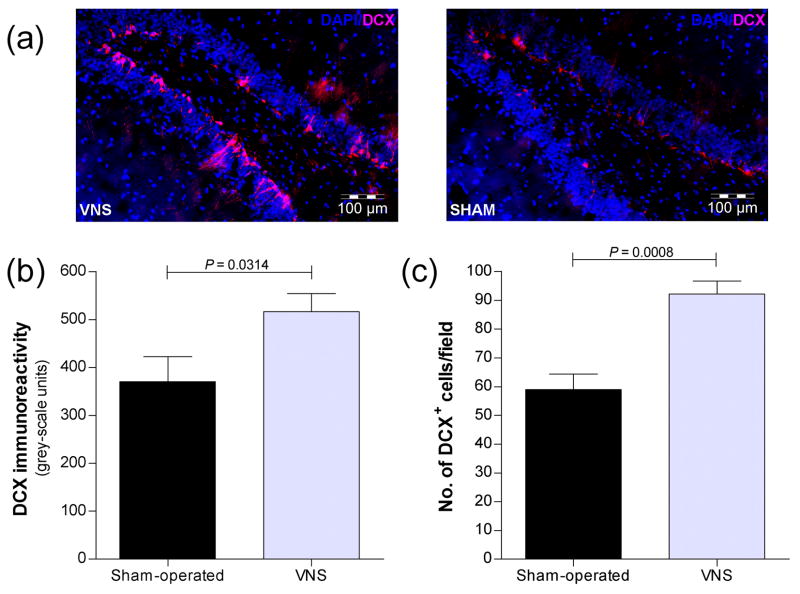
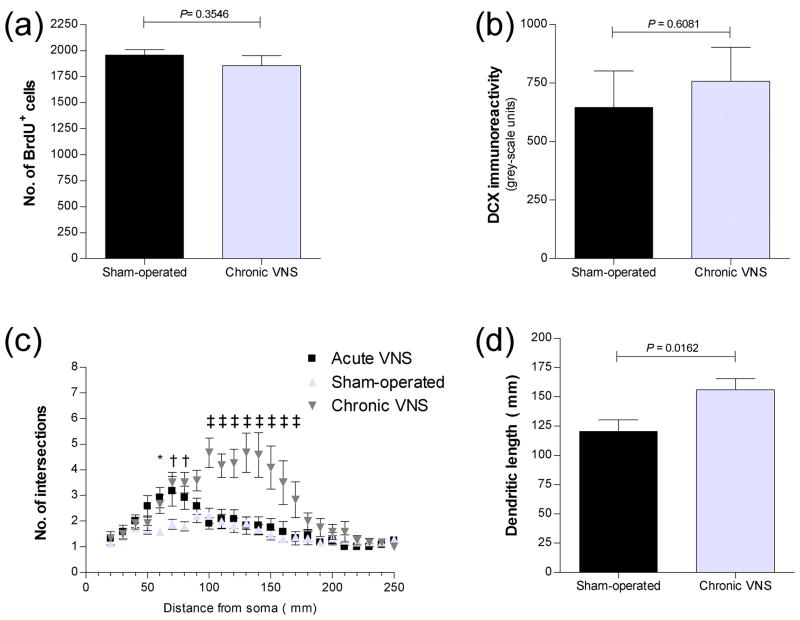
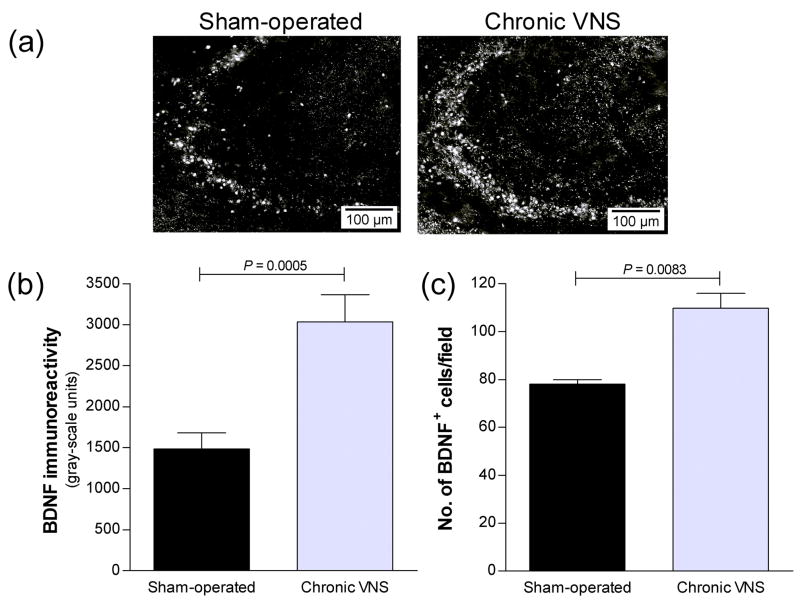
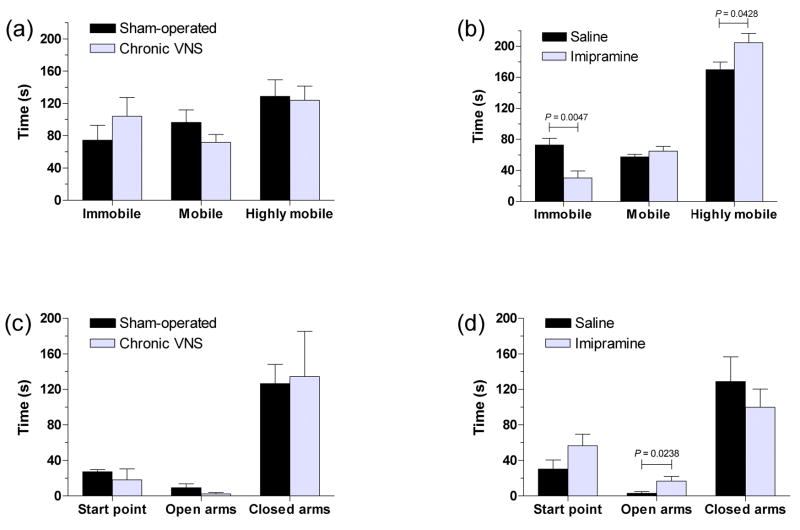
Vagus nerve stimulation (VNS) is used to treat pharmacotherapy-resistant epilepsy and depression. However, the mechanisms underlying the therapeutic efficacy of VNS remain unclear. We examined the effects of VNS on hippocampal neuronal plasticity and behaviour in rats. Cell proliferation in the hippocampus of rats subjected to acute (3 h) or chronic (1 month) VNS was examined by injection of bromodeoxyuridine (BrdU) and immunohistochemistry. Expression of doublecortin (DCX) and brain-derived neurotrophic factor (BDNF) was evaluated by immunofluorescence staining. The dendritic morphology of DCX+ neurons was measured by Sholl analysis. Our results show that acute VNS induced an increase in the number of BrdU+ cells in the dentate gyrus that was apparent 24 h and 3 wk after treatment. It also induced long-lasting increases in the amount of DCX immunoreactivity and in the number of DCX+ neurons. Neither the number of BrdU+ cells nor the amount of DCX immunoreactivity was increased 3 wk after the cessation of chronic VNS. Chronic VNS induced long-lasting increases in the amount of BDNF immunoreactivity and the number of BDNF+ cells as well as in the dendritic complexity of DCX+ neurons in the hippocampus. In contrast to chronic imipramine treatment, chronic VNS had no effect on the behaviour of rats in the forced swim or elevated plus-maze tests. Both chronic and acute VNS induced persistent changes in hippocampal neurons that may play a key role in the therapeutic efficacy of VNS. However, these changes were not associated with evident behavioural alterations characteristic of an antidepressant or anxiolytic action.
Conflict of interest statement
None
Figures
References
-
- Ben-Menachem E. Vagus-nerve stimulation for the treatment of epilepsy. Lancet Neurology. 2002;1(8):477–482. - PubMed
-
- Ben-Menachem E, Hamberger A, Hedner T, Hammond EJ, Uthman BM, Slater J, Treig T, Stefan H, Ramsay RE, Wernicke JF, et al. Effects of vagus nerve stimulation on amino acids and other metabolites in the CSF of patients with partial seizures. Epilepsy Research. 1995;20(3):221–227. - PubMed
-
- Ben-Menachem E, Manon-Espaillat R, Ristanovic R, Wilder BJ, Stefan H, Mirza W, Tarver WB, Wernicke JF. Vagus nerve stimulation for treatment of partial seizures: 1. A controlled study of effect on seizures. First International Vagus Nerve Stimulation Study Group. Epilepsia. 1994;35(3):616–626. - PubMed
-
- Bohning DE, Lomarev MP, Denslow S, Nahas Z, Shastri A, George MS. Feasibility of vagus nerve stimulation-synchronized blood oxygenation level-dependent functional MRI. Investigative Radiology. 2001;36(8):470–479. - PubMed
-
- Bruel-Jungerman E, Rampon C, Laroche S. Adult hippocampal neurogenesis, synaptic plasticity and memory: facts and hypotheses. Reviews in the Neurosciences. 2007;18(2):93–114. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
