

A dynamic stochastic model of frequency-dependent stress fiber alignment induced by cyclic stretch
- PMID: 19319193
- PMCID: PMC2656616
- DOI: 10.1371/journal.pone.0004853
A dynamic stochastic model of frequency-dependent stress fiber alignment induced by cyclic stretch
Abstract
Background: Actin stress fibers (SFs) are mechanosensitive structural elements that respond to forces to affect cell morphology, migration, signal transduction and cell function. Cells are internally stressed so that SFs are extended beyond their unloaded lengths, and SFs tend to self-adjust to an equilibrium level of extension. While there is much evidence that cells reorganize their SFs in response to matrix deformations, it is unclear how cells and their SFs determine their specific response to particular spatiotemporal changes in the matrix.
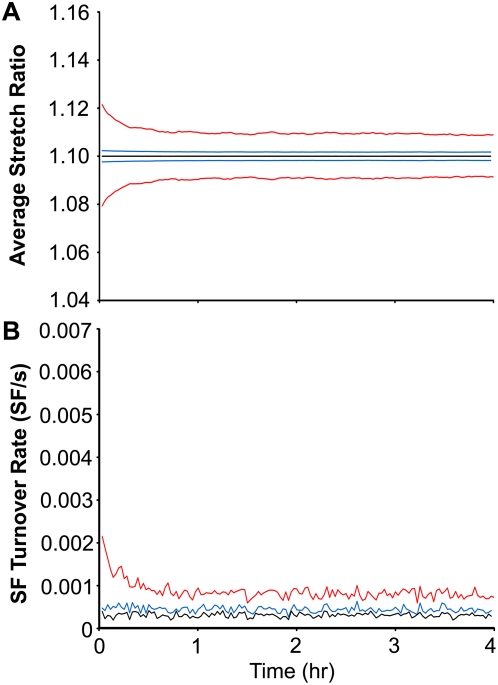
Methodology/principal findings: Bovine aortic endothelial cells were subjected to cyclic uniaxial stretch over a range of frequencies to quantify the rate and extent of stress fiber alignment. At a frequency of 1 Hz, SFs predominantly oriented perpendicular to stretch, while at 0.1 Hz the extent of SF alignment was markedly reduced and at 0.01 Hz there was no alignment at all. The results were interpreted using a simple kinematic model of SF networks in which the dynamic response depended on the rates of matrix stretching, SF turnover, and SF self-adjustment of extension. For these cells, the model predicted a threshold frequency of 0.01 Hz below which SFs no longer respond to matrix stretch, and a saturation frequency of 1 Hz above which no additional SF alignment would occur. The model also accurately described the dependence of SF alignment on matrix stretch magnitude.
Conclusions: The dynamic stochastic model was capable of describing SF reorganization in response to diverse temporal and spatial patterns of stretch. The model predicted that at high frequencies, SFs preferentially disassembled in the direction of stretch and achieved a new equilibrium by accumulating in the direction of lowest stretch. At low stretch frequencies, SFs self-adjusted to dissipate the effects of matrix stretch. Thus, SF turnover and self-adjustment are each important mechanisms that cells use to maintain mechanical homeostasis.
Conflict of interest statement
Figures





References
-
- Burridge K, Fath K, Kelly T, Nuckolls G, Turner C. Focal adhesions: transmembrane junctions between the extracellular matrix and the cytoskeleton. Annu Rev Cell Biol. 1988;4:487–525. - PubMed
-
- Deguchi S, Ohashi T, Sato M. Evaluation of tension in actin bundle of endothelial cells based on preexisting strain and tensile properties measurements. Mol Cell Biomech. 2005;2:125–133. - PubMed
-
- Lu L, Feng Y, Hucker WJ, Oswald SJ, Longmore GD, et al. Actin stress fiber pre-extension in human aortic endothelial cells. Cell Motil Cytoskeleton. 2008;65:281–294. - PubMed
-
- Lauffenburger DA, Horwitz AF. Cell migration: a physically integrated molecular process. Cell. 1996;84:359–369. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
