

Nramp1 promotes efficient macrophage recycling of iron following erythrophagocytosis in vivo
- PMID: 19321419
- PMCID: PMC2667064
- DOI: 10.1073/pnas.0900808106
Nramp1 promotes efficient macrophage recycling of iron following erythrophagocytosis in vivo
Abstract
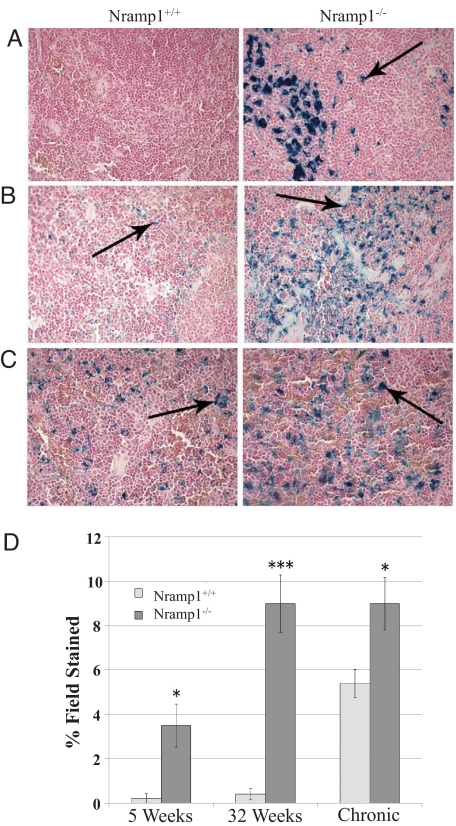
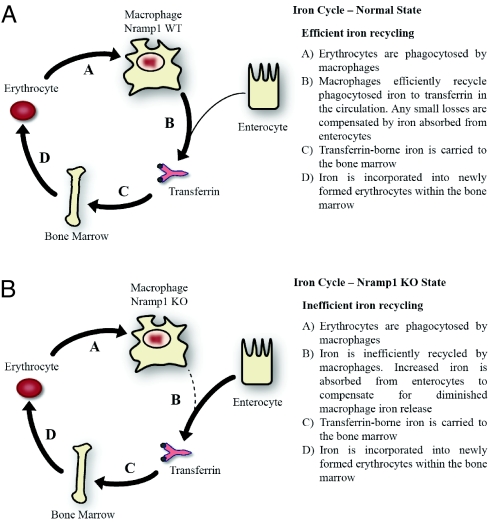
Natural resistance-associated macrophage protein 1 (Nramp1) is a divalent metal transporter expressed exclusively in phagocytic cells. We hypothesized that macrophage Nramp1 may participate in the recycling of iron acquired from phagocytosed senescent erythrocytes. To evaluate the role of Nramp1 in vivo, the iron parameters of WT and KO mice were analyzed after acute and chronic induction of hemolytic anemia. We found that untreated KO mice exhibited greater serum transferrin saturation and splenic iron content with higher duodenal ferroportin (Fpn) and divalent metal transporter 1 (DMT1) expression. Furthermore, hepatocyte iron content and hepcidin mRNA levels were dramatically lower in KO mice, indicating that hepcidin levels can be regulated by low-hepatocyte iron stores despite increased transferrin saturation. After acute treatment with the hemolytic agent phenylhydrazine (Phz), KO mice experienced a significant decrease in transferrin saturation and hematocrit, whereas WT mice were relatively unaffected. After a month-long Phz regimen, KO mice retained markedly increased quantities of iron within the liver and spleen and exhibited more pronounced splenomegaly and reticulocytosis than WT mice. After injection of (59)Fe-labeled heat-damaged reticulocytes, KO animals accumulated erythrophagocytosed (59)Fe within their liver and spleen, whereas WT animals efficiently recycled phagocytosed (59)Fe to the marrow and erythrocytes. These data imply that without Nramp1, iron accumulates within the liver and spleen during erythrophagocytosis and hemolytic anemia, supporting our hypothesis that Nramp1 promotes efficient hemoglobin iron recycling in macrophages. Our observations suggest that mutations in Nramp1 could result in a novel form of human hereditary iron overload.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Koury MJ, Ponka P. New insights into erythropoiesis: The roles of folate, vitamin B12, and iron. Annu Rev Nutr. 2004;24:105–131. - PubMed
-
- Sheftel AD, Kim SF, Ponka P. Non-heme induction of heme oxygenase-1 does not alter cellular iron metabolism. J Biol Chem. 2007;282:10480–10486. - PubMed
-
- Hershko C. Storage iron regulation. Prog Hematol. 1977;10:105–148. - PubMed
-
- Bothwell TH, Charlton RW, Cook JD, Finch CA. Iron Metabolism in Man. Oxford, UK: Blackwell Scientific; 1979.
-
- Ponka P. Cellular iron metabolism. Kidney Int Suppl. 1999;69:S2–S11. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
