

MDM2 acts downstream of p53 as an E3 ligase to promote FOXO ubiquitination and degradation
- PMID: 19321440
- PMCID: PMC2682847
- DOI: 10.1074/jbc.M901758200
MDM2 acts downstream of p53 as an E3 ligase to promote FOXO ubiquitination and degradation
Abstract
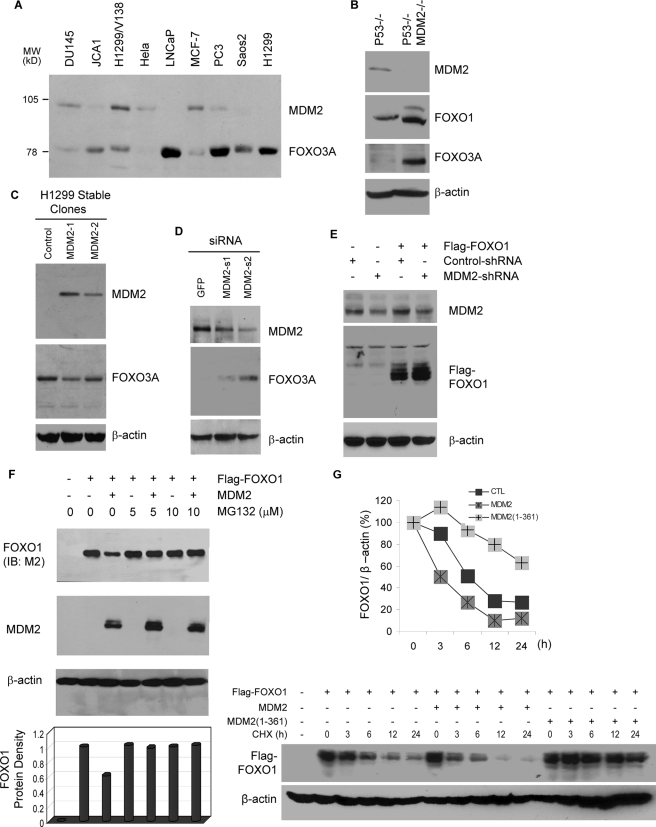
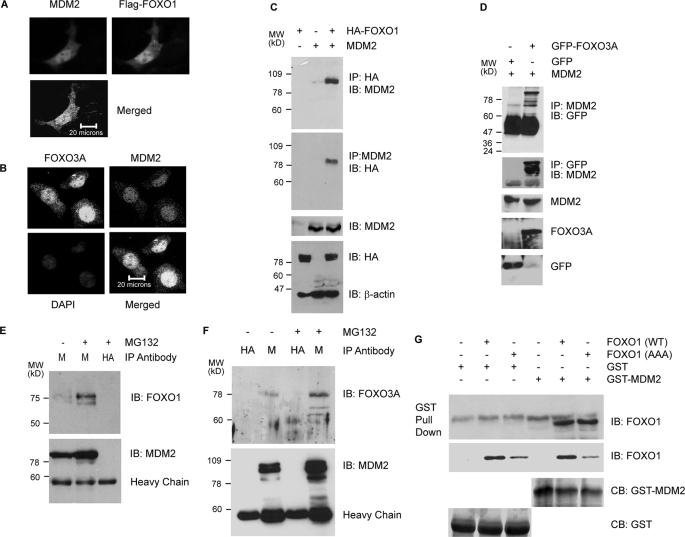
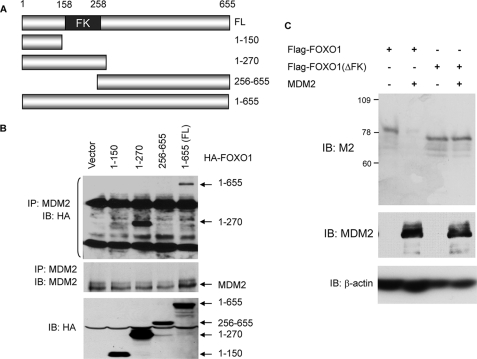
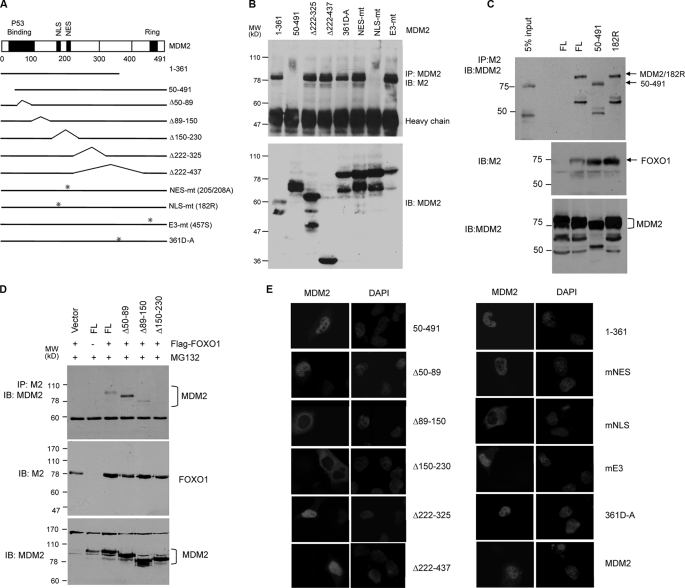
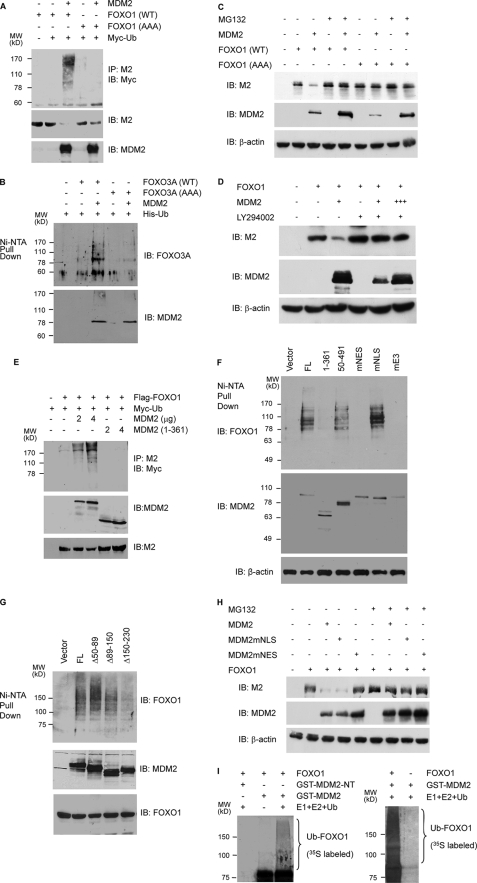
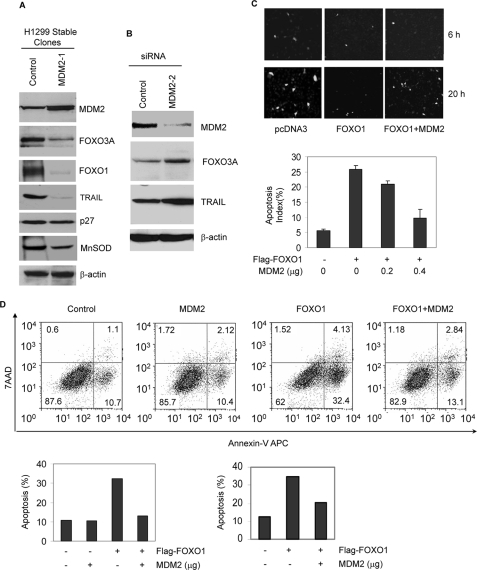
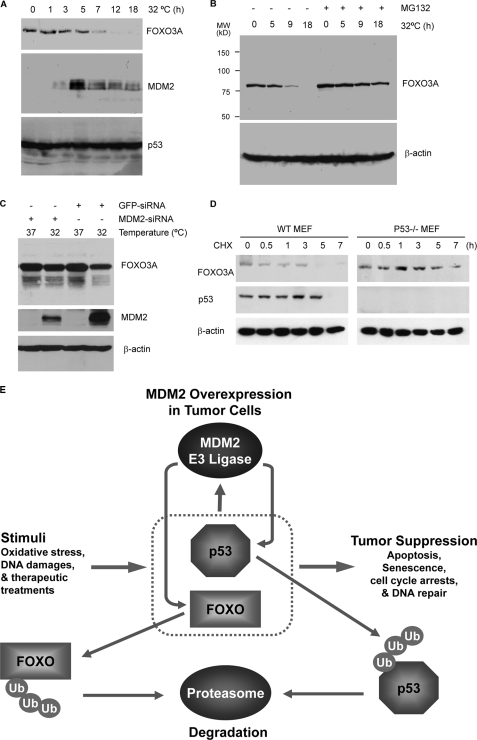
Members of the FOXO (forkhead O) class of transcription factors are tumor suppressors that also control aging and organismal life span. Mammalian FOXO degradation is proteasome-mediated, although the ubiquitin E3 ligase for FOXO factors remains to be defined. We show that MDM2 binds to FOXO1 and FOXO3A and promotes their ubiquitination and degradation, a process apparently dependent on FOXO phosphorylation at AKT sites and the E3 ligase activity of MDM2. Binding of MDM2 to FOXO occurs through the region of MDM2 that directs its cellular trafficking and the forkhead box of FOXO1. MDM2 promotes the ubiquitination of FOXO1 in a cell-free system, and its knockdown by small interfering RNA causes accumulation of endogenous FOXO3A protein in cells and enhances the expression of FOXO target genes. In cells stably expressing a temperature-sensitive p53 mutant, activation of p53 by shifting to permissive temperatures leads to MDM2 induction and degradation of endogenous FOXO3A. These data suggest that MDM2 acts as an ubiquitin E3 ligase, downstream of p53, to regulate the degradation of mammalian FOXO factors.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
