

Excitatory actions of ventral root stimulation during network activity generated by the disinhibited neonatal mouse spinal cord
- PMID: 19321640
- PMCID: PMC2694107
- DOI: 10.1152/jn.90740.2008
Excitatory actions of ventral root stimulation during network activity generated by the disinhibited neonatal mouse spinal cord
Abstract
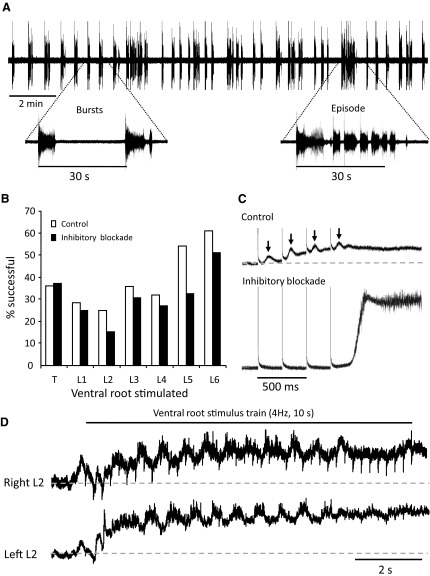
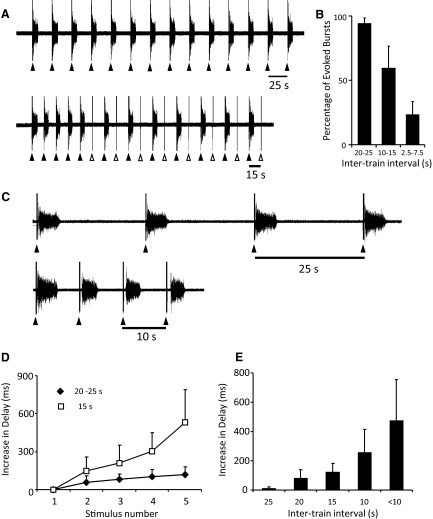
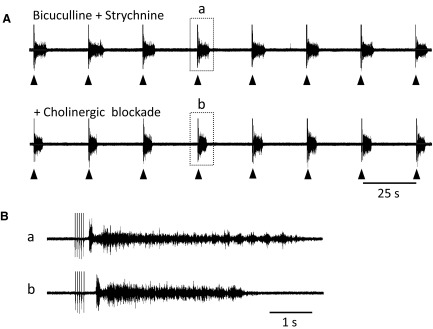
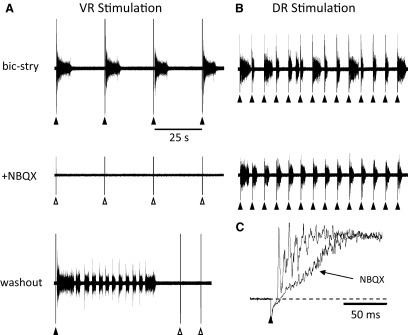
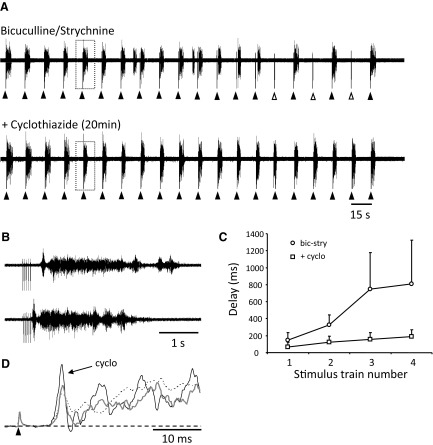
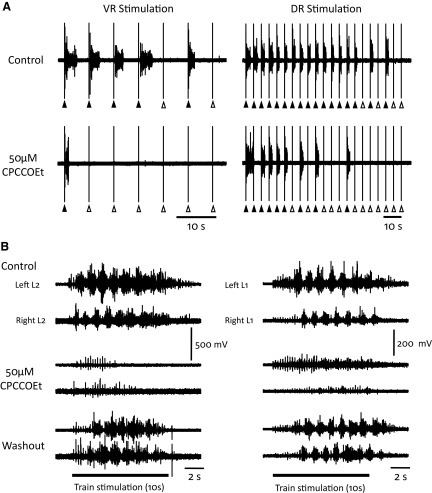
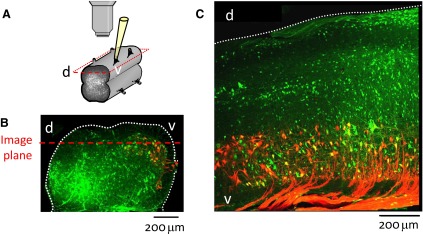
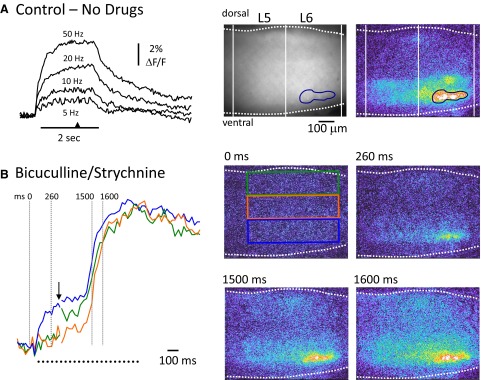
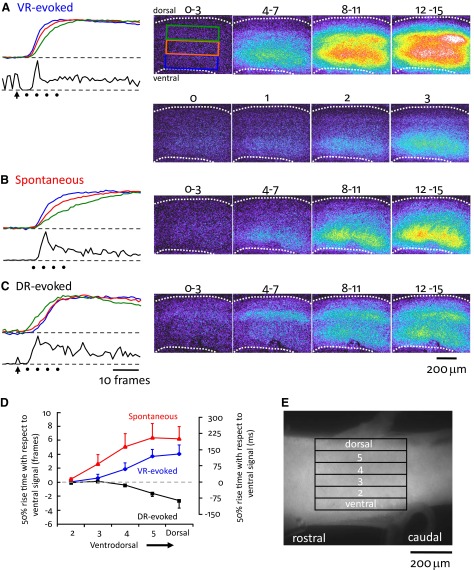
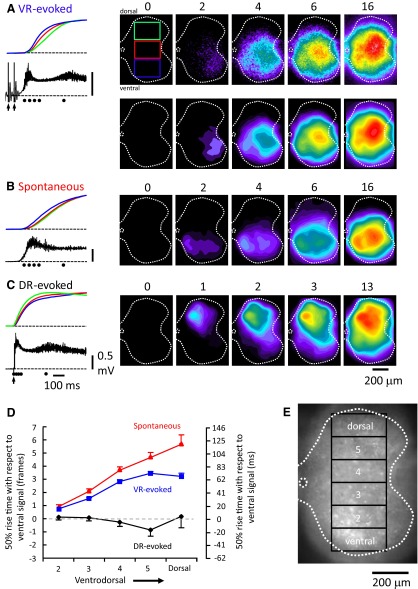
To further understand the excitatory effects of motoneurons on spinal network function, we investigated the entrainment of disinhibited rhythms by ventral root (VR) stimulation in the neonatal mouse spinal cord. A brief train of stimuli applied to a VR triggered bursting reliably in 31/32 experiments. The same roots that entrained disinhibited bursting could also produce locomotor-like activity with a similar probability when the network was not disinhibited. The ability of VR stimulation to entrain the rhythm persisted in nicotinic and muscarinic cholinergic antagonists but was blocked by the AMPAR antagonist NBQX. Bath application of the type I mGluR1 receptor antagonist CPCCOEt reduced the ability of both dorsal root and VR stimulation to entrain the disinhibited rhythm and abolished the ability of either type of stimulation to evoke locomotor-like activity. Calcium imaging through the lateral aspect of the cord revealed that VR stimulation and spontaneously occurring bursts were accompanied by a wave of activity that originated ventrally and propagated dorsally. Imaging the cut transverse face of L(5) revealed that the earliest VR-evoked optical activity began ventrolaterally. The optical activity accompanying spontaneous bursts could originate ventrolaterally, ventromedially, or throughout the mediolateral extent of the ventral horn or very occasionally dorsally. Collectively, our data indicate that VR stimulation can entrain disinhibited spinal network activity and trigger locomotor-like activity through a mechanism dependent on activation of both ionotropic and metabotropic glutamate receptors. The effects of entrainment appear to be mediated by a ventrolaterally located network that is also active during spontaneously occurring bursts.
Figures
Similar articles
-
Crossed rhythmic synaptic input to motoneurons during selective activation of the contralateral spinal locomotor network.J Neurosci. 1997 Dec 15;17(24):9433-47. doi: 10.1523/JNEUROSCI.17-24-09433.1997. J Neurosci. 1997. PMID: 9390999 Free PMC article.
-
Localization of rhythmogenic networks responsible for spontaneous bursts induced by strychnine and bicuculline in the rat isolated spinal cord.J Neurosci. 1996 Nov 1;16(21):7063-76. doi: 10.1523/JNEUROSCI.16-21-07063.1996. J Neurosci. 1996. PMID: 8824342 Free PMC article.
-
Mechanisms of excitation of spinal networks by stimulation of the ventral roots.Ann N Y Acad Sci. 2010 Jun;1198:63-71. doi: 10.1111/j.1749-6632.2010.05535.x. Ann N Y Acad Sci. 2010. PMID: 20536921 Free PMC article.
-
Spontaneous rhythmic bursts induced by pharmacological block of inhibition in lumbar motoneurons of the neonatal rat spinal cord.J Neurophysiol. 1996 Feb;75(2):640-7. doi: 10.1152/jn.1996.75.2.640. J Neurophysiol. 1996. PMID: 8714641
-
Rhythmogenic networks are potently modulated by activation of muscarinic acetylcholine receptors in the rodent spinal cord.J Neurochem. 2021 Sep;158(6):1263-1273. doi: 10.1111/jnc.15354. Epub 2021 Apr 12. J Neurochem. 2021. PMID: 33735482 Review.
Cited by
-
Sensory-Derived Glutamate Regulates Presynaptic Inhibitory Terminals in Mouse Spinal Cord.Neuron. 2016 Jun 15;90(6):1189-1202. doi: 10.1016/j.neuron.2016.05.008. Epub 2016 Jun 2. Neuron. 2016. PMID: 27263971 Free PMC article.
-
Motoneuronal Regulation of Central Pattern Generator and Network Function.Adv Neurobiol. 2022;28:259-280. doi: 10.1007/978-3-031-07167-6_11. Adv Neurobiol. 2022. PMID: 36066829
-
Properties of the epileptiform activity in the cingulate cortex of a mouse model of LIS1 dysfunction.Brain Struct Funct. 2022 Jun;227(5):1599-1614. doi: 10.1007/s00429-022-02458-1. Epub 2022 Feb 1. Brain Struct Funct. 2022. PMID: 35103859 Free PMC article.
-
Ventral root evoked entrainment of disinhibited bursts across early postnatal development in mice.IBRO Rep. 2020 Oct 27;9:310-318. doi: 10.1016/j.ibror.2020.10.005. eCollection 2020 Dec. IBRO Rep. 2020. PMID: 33294722 Free PMC article.
-
Expressions of VGLUT1/2 in the inspiratory interneurons and GAD65/67 in the inspiratory Renshaw cells in the neonatal rat upper thoracic spinal cord.IBRO Rep. 2018 Aug 4;5:24-32. doi: 10.1016/j.ibror.2018.08.001. eCollection 2018 Dec. IBRO Rep. 2018. PMID: 30135953 Free PMC article.
References
-
- Beato M, Nistri A. Interaction between disinhibited bursting and fictive locomotor patterns in the rat isolated spinal cord. J Neurophysiol. 82: 2029–2038, 1999. - PubMed
-
- Biscoe TJ, Nickels SM, Stirling CA. Numbers and sizes of nerve fibres in mouse spinal roots. Q J Exp Physiol 67: 473–494, 1982. - PubMed
-
- Bonnot A, Mentis GZ, Skoch J, O'Donovan MJ. Electroporation loading of calcium-sensitive dyes into the central nervous system J Neurophysiol 93: 1793–1808, 2005. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources