

Trf1 is not required for proliferation or functional telomere maintenance in chicken DT40 cells
- PMID: 19321665
- PMCID: PMC2682597
- DOI: 10.1091/mbc.e08-10-1019
Trf1 is not required for proliferation or functional telomere maintenance in chicken DT40 cells
Abstract
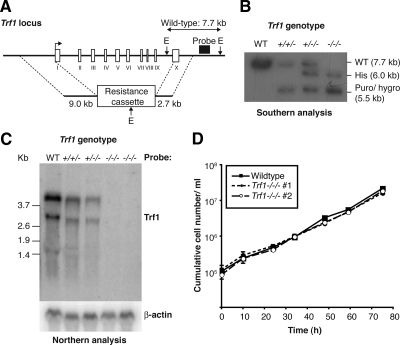
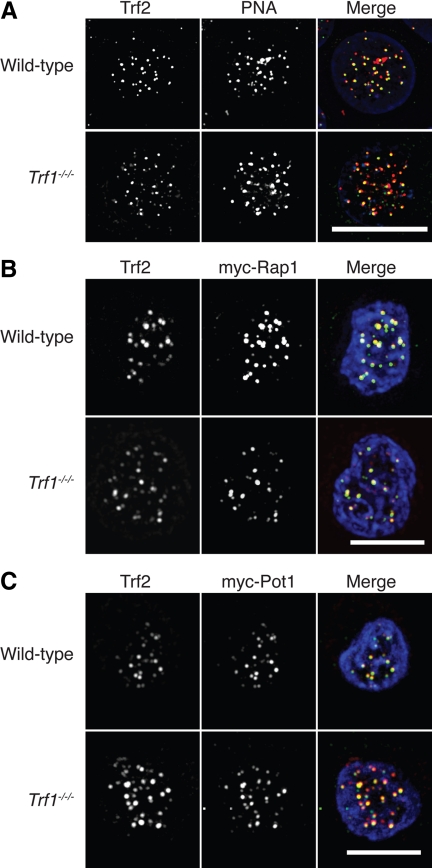
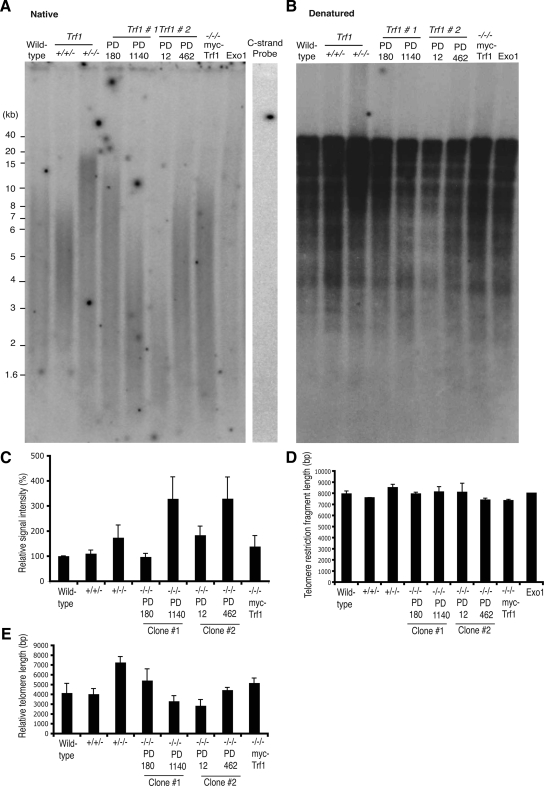
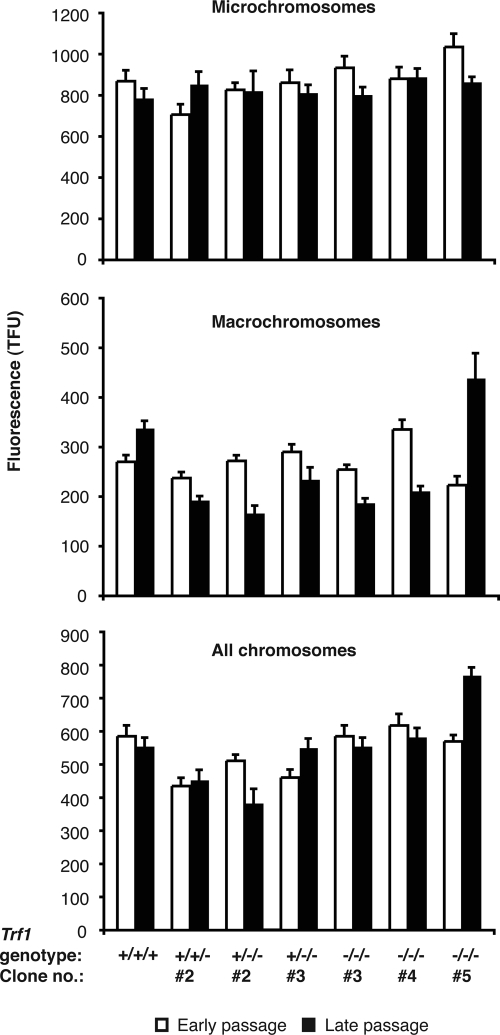
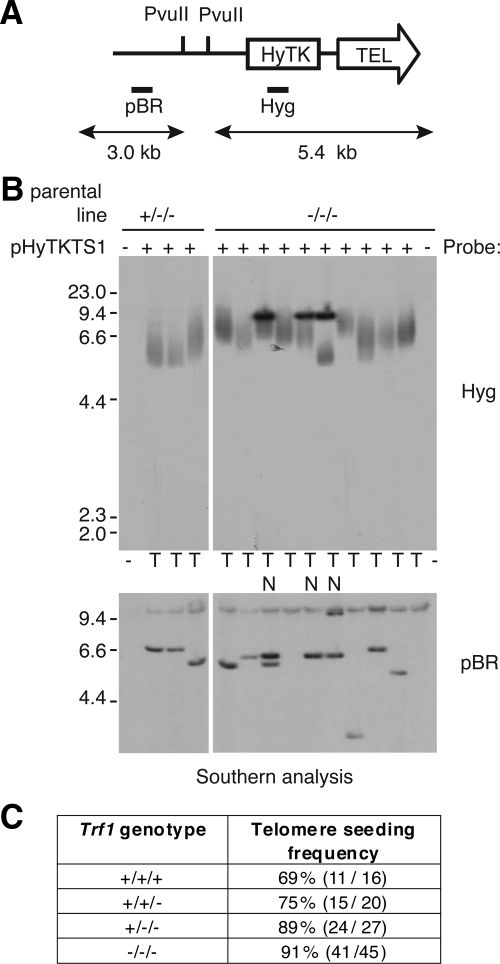
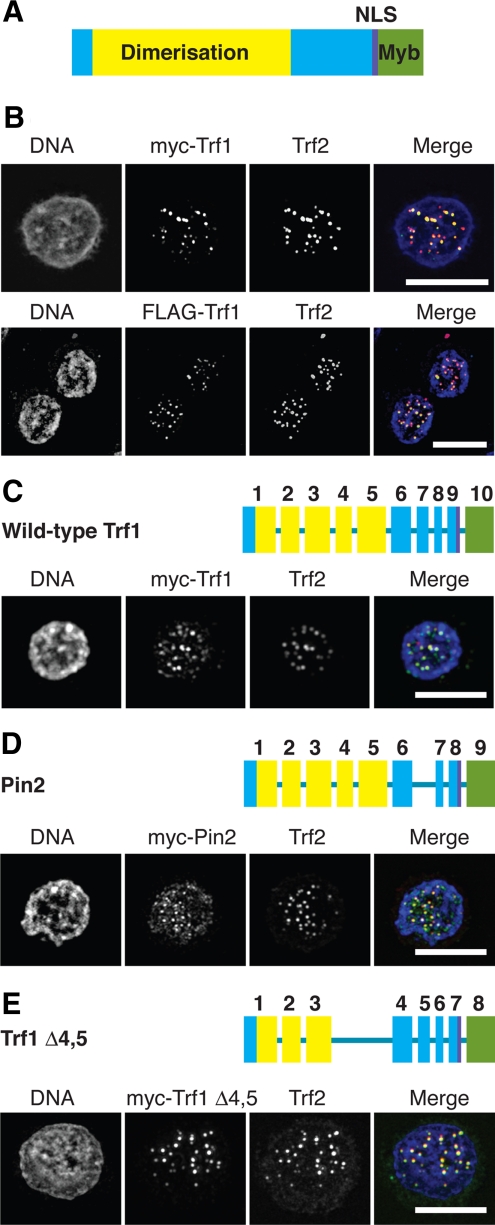
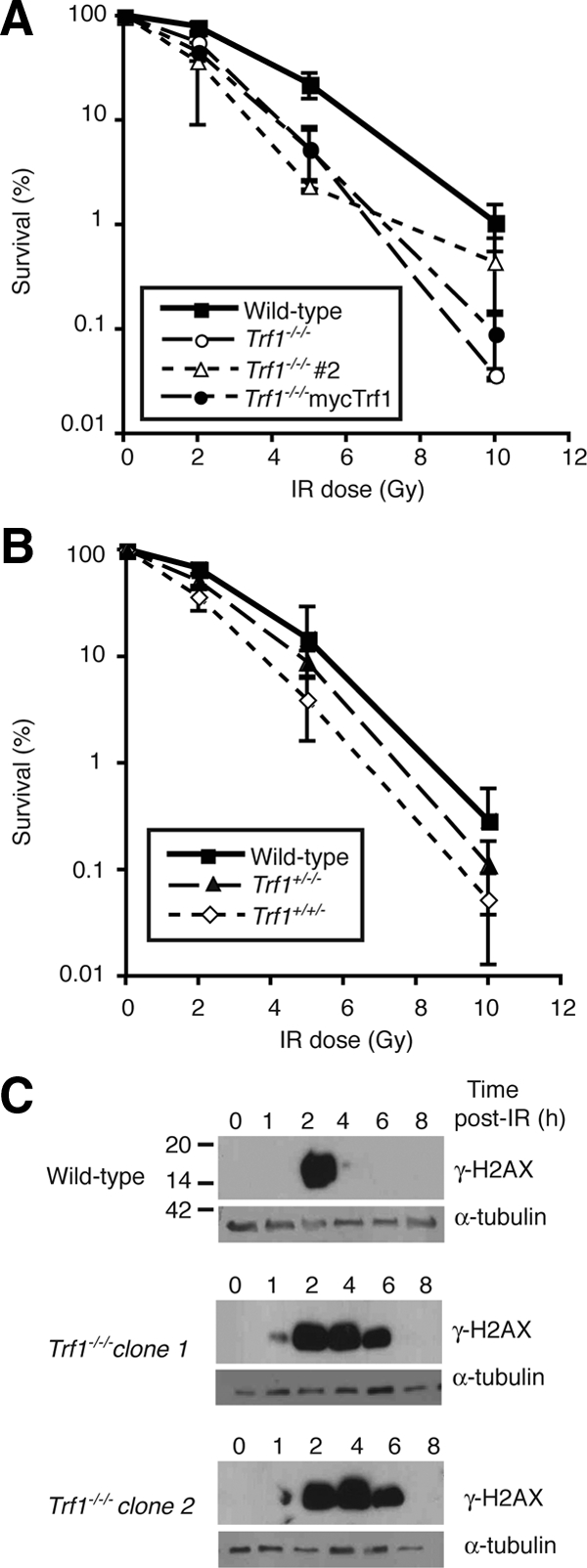
The telomere end-protection complex prevents the ends of linear eukaryotic chromosomes from degradation or inappropriate DNA repair. The homodimeric double-stranded DNA-binding protein, Trf1, is a component of this complex and is essential for mouse embryonic development. To define the requirement for Trf1 in somatic cells, we deleted Trf1 in chicken DT40 cells by gene targeting. Trf1-deficient cells proliferated as rapidly as control cells and showed telomeric localization of Trf2, Rap1, and Pot1. Telomeric G-strand overhang lengths were increased in late-passage Trf1-deficient cells, although telomere lengths were unaffected by Trf1 deficiency, as determined by denaturing Southern and quantitative FISH analysis. Although we observed some clonal variation in terminal telomere fragment lengths, this did not correlate with cellular Trf1 levels. Trf1 was not required for telomere seeding, indicating that de novo telomere formation can proceed without Trf1. The Pin2 isoform and a novel exon 4, 5-deleted isoform localized to telomeres in Trf1-deficient cells. Trf1-deficient cells were sensitive to DNA damage induced by ionizing radiation. Our data demonstrate that chicken DT40 B cells do not require Trf1 for functional telomere structure and suggest that Trf1 may have additional, nontelomeric roles involved in maintaining genome stability.
Figures
References
-
- Bailey S. M., Brenneman M. A., Halbrook J., Nickoloff J. A., Ullrich R. L., Goodwin E. H. The kinase activity of DNA-PK is required to protect mammalian telomeres. DNA Repair. 2004;3:225–233. - PubMed
-
- Bianchi A., de Lange T. Ku binds telomeric DNA in vitro. J. Biol. Chem. 1999;274:21223–21227. - PubMed
-
- Blackburn E. H. Switching and signaling at the telomere. Cell. 2001;106:661–673. - PubMed
-
- Bradshaw P. S., Stavropoulos D. J., Meyn M. S. Human telomeric protein TRF2 associates with genomic double-strand breaks as an early response to DNA damage. Nat. Genet. 2005;37:193–197. - PubMed
-
- Celli G. B., de Lange T. DNA processing is not required for ATM-mediated telomere damage response after TRF2 deletion. Nat. Cell Biol. 2005;7:712–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
