

Ubiquitin-mediated proteolysis of HuR by heat shock
- PMID: 19322201
- PMCID: PMC2683047
- DOI: 10.1038/emboj.2009.67
Ubiquitin-mediated proteolysis of HuR by heat shock
Abstract
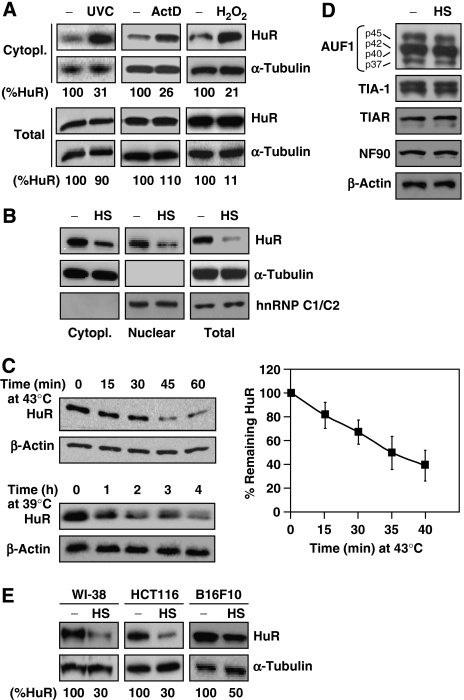
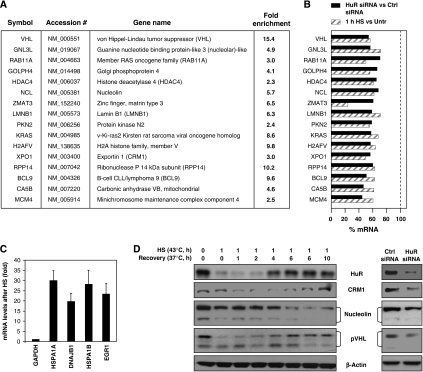
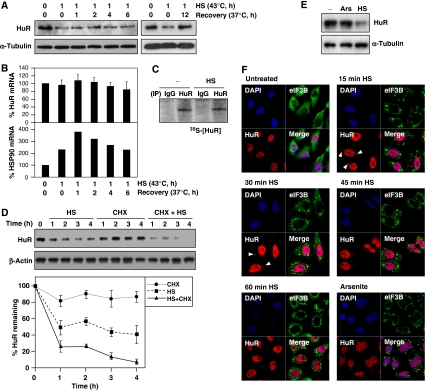
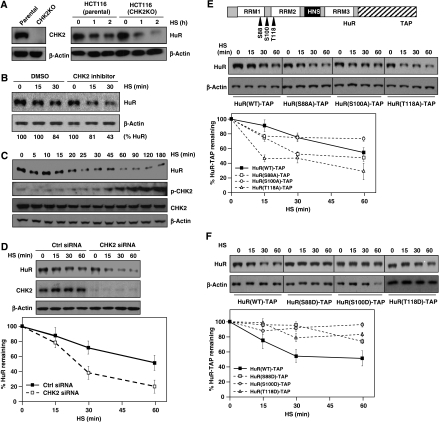
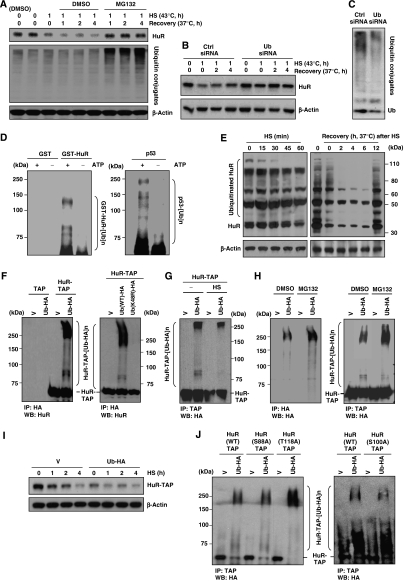
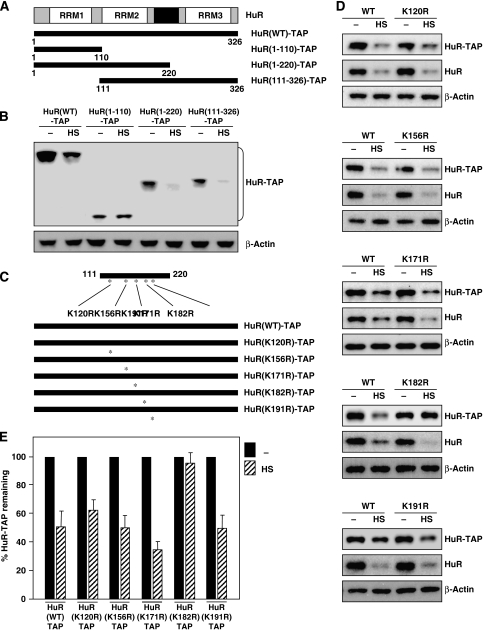
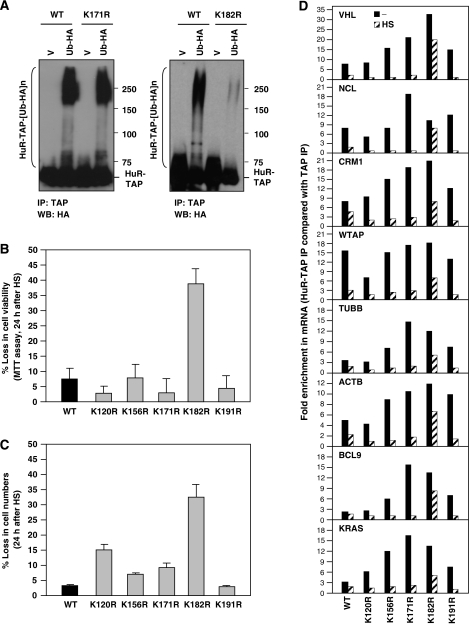
The RNA-binding protein HuR regulates the stability and translation of numerous mRNAs encoding stress-response and proliferative proteins. Although its post-transcriptional influence has been linked primarily to its cytoplasmic translocation, here we report that moderate heat shock (HS) potently reduces HuR levels, thereby altering the expression of HuR target mRNAs. HS did not change HuR mRNA levels or de novo translation, but instead reduced HuR protein stability. Supporting the involvement of the ubiquitin-proteasome system in this process were results showing that (1) HuR was ubiquitinated in vitro and in intact cells, (2) proteasome inhibition increased HuR abundance after HS, and (3) the HuR kinase checkpoint kinase 2 protected against the loss of HuR by HS. Within a central, HS-labile approximately 110-amino-acid region, K182 was found to be essential for HuR ubiquitination and proteolysis as mutant HuR(K182R) was left virtually unubiquitinated and was refractory to HS-triggered degradation. Our findings reveal that HS transiently lowers HuR by proteolysis linked to K182 ubiquitination and that HuR reduction enhances cell survival following HS.
Figures
References
-
- Abdelmohsen K, Lal A, Kim HH, Gorospe M (2007b) Posttranscriptional orchestration of an anti-apoptotic program by HuR. Cell Cycle 6: 1288–1292 - PubMed
-
- Bhattacharyya SN, Habermacher R, Martine U, Closs EI, Filipowicz W (2006) Relief of microRNA-mediated translational repression in human cells subjected to stress. Cell 125: 1111–1124 - PubMed
-
- Bonelli MA, Alfieri RR, Desenzani S, Petronini PG, Borghetti AF (2004) Proteasome inhibition increases HuR level, restores heat-inducible HSP72 expression and thermotolerance in WI-38 senescent human fibroblasts. Exp Gerontol 39: 423–432 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
