

Presynaptic regulation of astroglial excitatory neurotransmitter transporter GLT1
- PMID: 19323997
- PMCID: PMC2743171
- DOI: 10.1016/j.neuron.2009.02.010
Presynaptic regulation of astroglial excitatory neurotransmitter transporter GLT1
Abstract
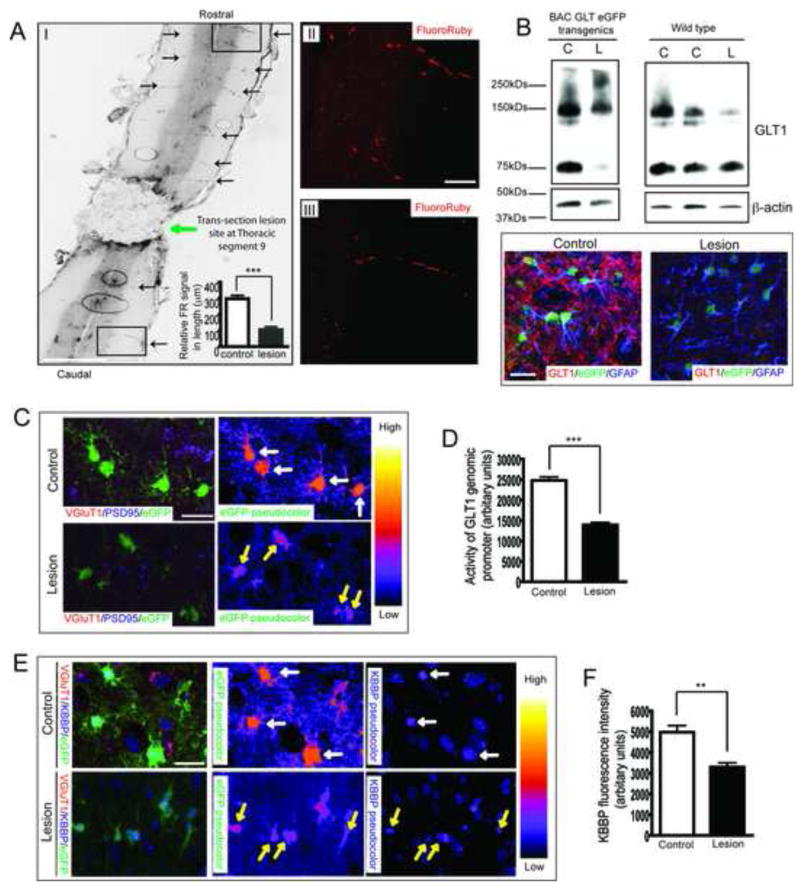
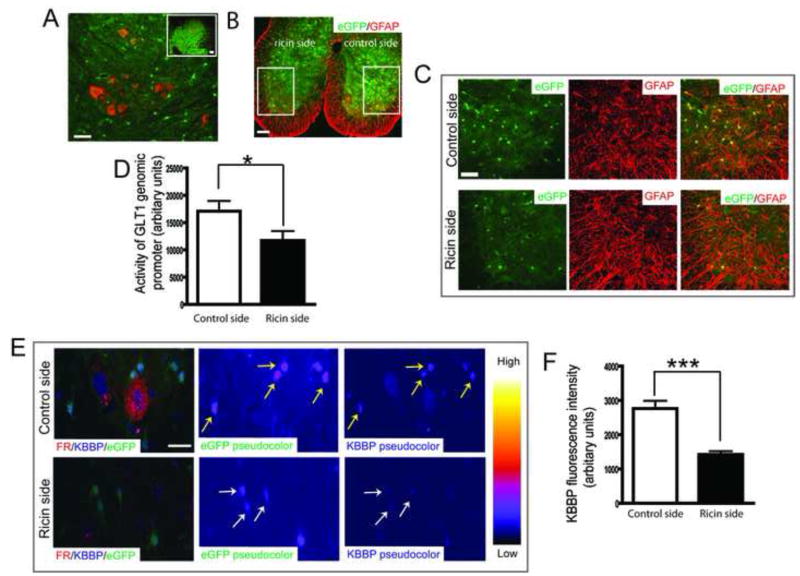
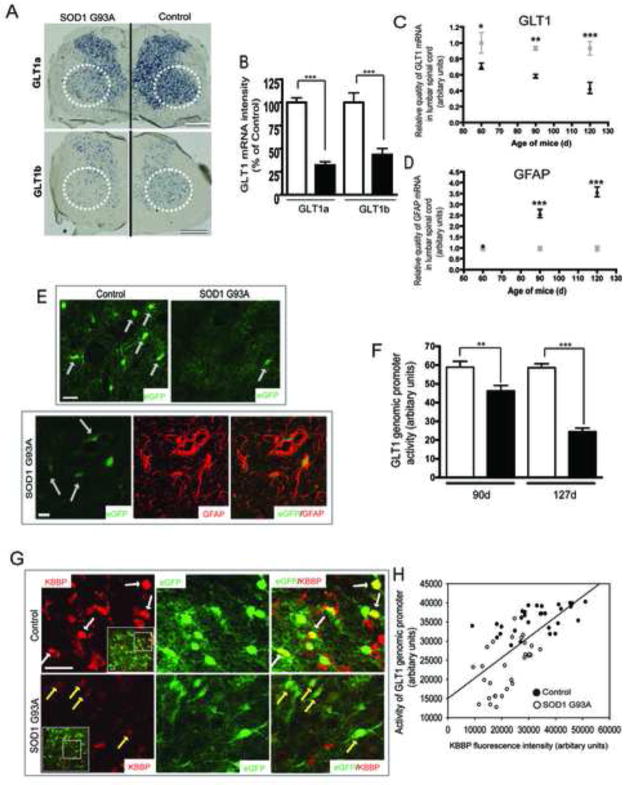
The neuron-astrocyte synaptic complex is a fundamental operational unit of the nervous system. Astroglia regulate synaptic glutamate, via neurotransmitter transport by GLT1/EAAT2. Astroglial mechanisms underlying this essential neuron-glial communication are not known. We now show that presynaptic terminals regulate astroglial synaptic functions, GLT1/EAAT2, via kappa B-motif binding phosphoprotein (KBBP), the mouse homolog of human heterogeneous nuclear ribonucleoprotein K (hnRNP K), which binds the GLT1/EAAT2 promoter. Neuron-stimulated KBBP is required for GLT1/EAAT2 transcriptional activation and is responsible for astroglial alterations in neural injury. Denervation of neuron-astrocyte signaling by corticospinal tract transection, ricin-induced motor neuron death, or neurodegeneration in amyotrophic lateral sclerosis all result in reduced astroglial KBBP expression and transcriptional dysfunction of astroglial transporter expression. Presynaptic elements dynamically coordinate normal astroglial function and also provide a fundamental signaling mechanism by which altered neuronal function and injury leads to dysregulated astroglia in CNS disease.
Figures
Comment in
-
What the neuron tells glia.Neuron. 2009 Mar 26;61(6):811-2. doi: 10.1016/j.neuron.2009.03.008. Neuron. 2009. PMID: 19323989
Similar articles
-
Epigenetic regulation of neuron-dependent induction of astroglial synaptic protein GLT1.Glia. 2010 Feb;58(3):277-86. doi: 10.1002/glia.20922. Glia. 2010. PMID: 19672971 Free PMC article.
-
Molecular comparison of GLT1+ and ALDH1L1+ astrocytes in vivo in astroglial reporter mice.Glia. 2011 Feb;59(2):200-7. doi: 10.1002/glia.21089. Glia. 2011. PMID: 21046559 Free PMC article.
-
Neuronal exosomal miRNA-dependent translational regulation of astroglial glutamate transporter GLT1.J Biol Chem. 2013 Mar 8;288(10):7105-16. doi: 10.1074/jbc.M112.410944. Epub 2013 Jan 30. J Biol Chem. 2013. PMID: 23364798 Free PMC article.
-
Role of excitatory amino acid transporter-2 (EAAT2) and glutamate in neurodegeneration: opportunities for developing novel therapeutics.J Cell Physiol. 2011 Oct;226(10):2484-93. doi: 10.1002/jcp.22609. J Cell Physiol. 2011. PMID: 21792905 Free PMC article. Review.
-
Morphological plasticity of astroglia: Understanding synaptic microenvironment.Glia. 2015 Dec;63(12):2133-51. doi: 10.1002/glia.22821. Epub 2015 Mar 18. Glia. 2015. PMID: 25782611 Free PMC article. Review.
Cited by
-
Transcriptional Regulation of Glutamate Transporters: From Extracellular Signals to Transcription Factors.Adv Pharmacol. 2016;76:103-45. doi: 10.1016/bs.apha.2016.01.004. Epub 2016 Mar 24. Adv Pharmacol. 2016. PMID: 27288076 Free PMC article. Review.
-
Rapid Regulation of Glutamate Transport: Where Do We Go from Here?Neurochem Res. 2022 Jan;47(1):61-84. doi: 10.1007/s11064-021-03329-7. Epub 2021 Apr 24. Neurochem Res. 2022. PMID: 33893911 Free PMC article. Review.
-
Exosome reporter mice reveal the involvement of exosomes in mediating neuron to astroglia communication in the CNS.Nat Commun. 2019 Sep 12;10(1):4136. doi: 10.1038/s41467-019-11534-w. Nat Commun. 2019. PMID: 31515491 Free PMC article.
-
Down-expressed GLT-1 in PSD astrocytes inhibits synaptic formation of NSC-derived neurons in vitro.Cell Cycle. 2019 Jan;18(1):105-114. doi: 10.1080/15384101.2018.1560201. Epub 2018 Dec 26. Cell Cycle. 2019. PMID: 30558468 Free PMC article.
-
Targeted over-expression of glutamate transporter 1 (GLT-1) reduces ischemic brain injury in a rat model of stroke.PLoS One. 2011;6(8):e22135. doi: 10.1371/journal.pone.0022135. Epub 2011 Aug 10. PLoS One. 2011. PMID: 21853027 Free PMC article.
References
-
- Arnth-Jensen N, Jabaudon D, Scanziani M. Cooperation between independent hippocampal synapses is controlled by glutamate uptake. Nat Neurosci. 2002;5:325–331. - PubMed
-
- Blinzinger K, Kreutzberg G. Displacement of synaptic terminals from regenerating motoneurons by microglial cells. Z Zellforsch Mikrosk Anat. 1968;85:145–157. - PubMed
-
- Bomsztyk K, Denisenko O, Ostrowski J. hnRNP K: one protein multiple processes. Bioessays. 2004;26:629–638. - PubMed
-
- Bristol LA, Rothstein JD. Glutamate transporter gene expression in amyotrophic lateral sclerosis motor cortex. Ann Neurol. 1996;39:676–679. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
