

Maternal effects in cooperative breeders: from hymenopterans to humans
- PMID: 19324618
- PMCID: PMC2666687
- DOI: 10.1098/rstb.2008.0298
Maternal effects in cooperative breeders: from hymenopterans to humans
Abstract
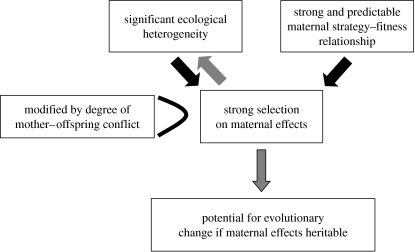
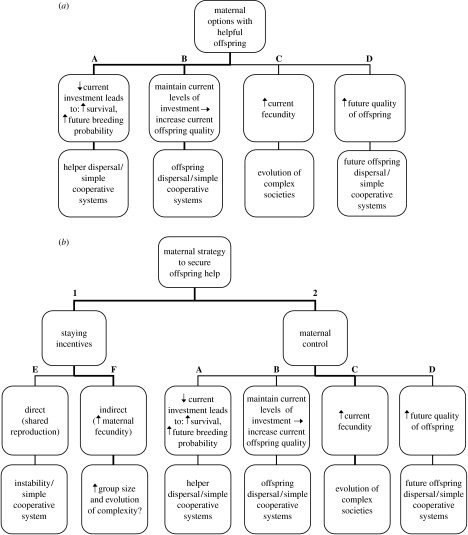
The environment that an offspring experiences during its development can have lifelong consequences for its morphology, anatomy, physiology and behaviour that are strong enough to span generations. One aspect of an offspring's environment that can have particularly pronounced and long-lasting effects is that provided by its parent(s) (maternal effects). Some disciplines in biology have been quicker to appreciate maternal effects than others, and some organisms provide better model systems for understanding the causes and consequences of the maternal environment for ecology and evolution than others. One field in which maternal effects has been poorly represented, and yet is likely to represent a particularly fruitful area for research, is the field of cooperative breeding (i.e. systems where offspring are reared by carers in addition to parent(s)). Here, we attempt to illustrate the scope of cooperative breeding systems for maternal effects research and, conversely, highlight the importance of maternal effects research for understanding cooperative breeding systems. To this end, we first outline why mothers will commonly benefit from affecting the phenotype of their offspring in cooperative breeding systems, present potential strategies that mothers could employ in order to do so and offer predictions regarding the circumstances under which different types of maternal effects might be expected. Second, we highlight why a neglect of maternal strategies and the effects that they have on their offspring could lead to miscalculations of helper/worker fitness gains and a misunderstanding of the factors selecting for the evolution and maintenance of cooperative breeding. Finally, we introduce the possibility that maternal effects could have significant consequences for our understanding of both the evolutionary origins of cooperative breeding and the rise of social complexity in cooperative systems.
Figures
Similar articles
-
Family living sets the stage for cooperative breeding and ecological resilience in birds.PLoS Biol. 2017 Jun 21;15(6):e2000483. doi: 10.1371/journal.pbio.2000483. eCollection 2017 Jun. PLoS Biol. 2017. PMID: 28636615 Free PMC article.
-
When mothers need others: The impact of hominin life history evolution on cooperative breeding.J Hum Evol. 2015 Jul;84:16-24. doi: 10.1016/j.jhevol.2015.01.009. Epub 2015 Apr 3. J Hum Evol. 2015. PMID: 25843884
-
Life histories and the evolution of cooperative breeding in mammals.Proc Biol Sci. 2012 Oct 7;279(1744):4065-70. doi: 10.1098/rspb.2012.1433. Epub 2012 Aug 8. Proc Biol Sci. 2012. PMID: 22874752 Free PMC article.
-
Variable postpartum responsiveness among humans and other primates with "cooperative breeding": A comparative and evolutionary perspective.Horm Behav. 2016 Jan;77:272-83. doi: 10.1016/j.yhbeh.2015.10.016. Epub 2015 Oct 28. Horm Behav. 2016. PMID: 26518662 Review.
-
Adult sex ratios and their implications for cooperative breeding in birds.Philos Trans R Soc Lond B Biol Sci. 2017 Sep 19;372(1729):20160322. doi: 10.1098/rstb.2016.0322. Philos Trans R Soc Lond B Biol Sci. 2017. PMID: 28760763 Free PMC article. Review.
Cited by
-
Maternal and cohort effects modulate offspring responses to multiple stressors.Proc Biol Sci. 2020 Jun 24;287(1929):20200492. doi: 10.1098/rspb.2020.0492. Epub 2020 Jun 17. Proc Biol Sci. 2020. PMID: 32546091 Free PMC article.
-
Parental manipulation of offspring size in social groups: a test using paper wasps.Behav Ecol Sociobiol. 2019;73(3):36. doi: 10.1007/s00265-019-2646-3. Epub 2019 Feb 25. Behav Ecol Sociobiol. 2019. PMID: 30880867 Free PMC article.
-
Maternal effects in relation to helper presence in the cooperatively breeding sociable weaver.PLoS One. 2013;8(3):e59336. doi: 10.1371/journal.pone.0059336. Epub 2013 Mar 25. PLoS One. 2013. PMID: 23536872 Free PMC article.
-
The evolution of early-life effects on social behaviour-why should social adversity carry over to the future?Philos Trans R Soc Lond B Biol Sci. 2019 Apr 15;374(1770):20180111. doi: 10.1098/rstb.2018.0111. Philos Trans R Soc Lond B Biol Sci. 2019. PMID: 30966877 Free PMC article.
-
A comparative study of an innate immune response in Lamprologine cichlid fishes.Naturwissenschaften. 2014 Oct;101(10):839-49. doi: 10.1007/s00114-014-1225-8. Epub 2014 Aug 19. Naturwissenschaften. 2014. PMID: 25135814
References
-
- Alexander R.D. The evolution of social behaviour. Annu. Rev. Ecol. Syst. 1974;5:325–383. doi:10.1146/annurev.es.05.110174.001545 - DOI
-
- Alexander R.D., Noonan K., Crespi B.J. The evolution of eusociality. In: Sherman P.W., Jarvis J., Alexander R.D., editors. The biology of the naked mole rat. Princeton University Press; Princeton, NJ: 1991. pp. 3–44.
-
- Anderson K.E., Linksvayer T.A., Smith C.R. The causes and consequences of genetic caste determination in ants (Hymenoptera: Formicidae) Myrmecol. News. 2008;11:119–132.
-
- Arnold K.E., Owens I.P.F. Cooperative breeding in birds: a comparative analysis of the life history hypothesis. Proc. R. Soc. B. 1998;265:739–745. doi:10.1098/rspb.1998.0355 - DOI
-
- Arnold K.E., Owens I.P.F., Goldizen A.W. Division of labour within cooperatively breeding groups. Behaviour. 2005;142:1577–1590. doi:10.1163/156853905774831927 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
