

Tat-binding protein-1 (TBP-1), an ATPase of 19S regulatory particles of the 26S proteasome, enhances androgen receptor function in cooperation with TBP-1-interacting protein/Hop2
- PMID: 19325002
- PMCID: PMC2703560
- DOI: 10.1210/en.2008-1122
Tat-binding protein-1 (TBP-1), an ATPase of 19S regulatory particles of the 26S proteasome, enhances androgen receptor function in cooperation with TBP-1-interacting protein/Hop2
Abstract
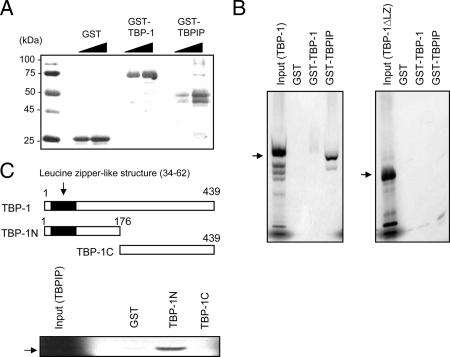
The 26S proteasome, which degrades ubiquitinated proteins, appears to contribute to the cyclical loading of androgen receptor (AR) to androgen response elements of target gene promoters; however, the mechanism whereby the 26S proteasome modulates AR recruitment remains unknown. Using yeast two-hybrid screening, we previously identified Tat-binding protein-1 (TBP-1), an adenosine triphosphatase of 19S regulatory particles of the 26S proteasome, as a transcriptional coactivator of thyroid hormone receptor. Independently, TBP-1-interacting protein (TBPIP) was also identified as a coactivator of several nuclear receptors, including AR. Here, we investigated whether TBP-1 could interact with and modulate transcriptional activation by AR cooperatively with TBPIP. TBP-1 mRNA was ubiquitously expressed in human tissues, including the testis and prostate, as well as in LNCaP cells. TBP-1 directly bound TBPIP through the amino-terminal domain possessing the leucine zipper structure. AR is physically associated with TBP-1 and TBPIP in vitro and in LNCaP cells. TBP-1 similarly and additively augmented AR-mediated transcription upon coexpression with TBPIP, and the ATPase domain, as well as leucine zipper structure in TBP-1, was essential for transcriptional enhancement. Overexpression of TBP-1 did not alter AR protein and mRNA levels. In the chromatin immunoprecipitation assay, TBP-1 was transiently recruited to the proximal androgen response element of the prostate-specific antigen gene promoter in a ligand-dependent manner in LNCaP cells. These findings suggest that a component of 19S regulatory particles directly binds AR and might participate in AR-mediated transcriptional activation in cooperation with TBPIP.
Figures






Similar articles
-
Molecular cloning and characterization of a novel TBP-1 interacting protein (TBPIP):enhancement of TBP-1 action on Tat by TBPIP.Biochem Biophys Res Commun. 1997 Oct 9;239(1):176-81. doi: 10.1006/bbrc.1997.7447. Biochem Biophys Res Commun. 1997. PMID: 9345291
-
Human immunodeficiency virus type 1 Tat binding protein-1 is a transcriptional coactivator specific for TR.Mol Endocrinol. 2001 Aug;15(8):1329-43. doi: 10.1210/mend.15.8.0680. Mol Endocrinol. 2001. PMID: 11463857
-
The role of hepatocyte nuclear factor-3 alpha (Forkhead Box A1) and androgen receptor in transcriptional regulation of prostatic genes.Mol Endocrinol. 2003 Aug;17(8):1484-507. doi: 10.1210/me.2003-0020. Epub 2003 May 15. Mol Endocrinol. 2003. PMID: 12750453
-
Expression and function of androgen receptor coactivators in prostate cancer.J Steroid Biochem Mol Biol. 2004 Nov;92(4):265-71. doi: 10.1016/j.jsbmb.2004.10.003. Epub 2004 Dec 19. J Steroid Biochem Mol Biol. 2004. PMID: 15663989 Review.
-
Regulatory processes affecting androgen receptor expression, stability, and function: potential targets to treat hormone-refractory prostate cancer.J Cell Biochem. 2006 Aug 15;98(6):1408-23. doi: 10.1002/jcb.20927. J Cell Biochem. 2006. PMID: 16619263 Review.
Cited by
-
Ubiquitin and proteasomes in transcription.Annu Rev Biochem. 2012;81:177-201. doi: 10.1146/annurev-biochem-052110-120012. Epub 2012 Mar 8. Annu Rev Biochem. 2012. PMID: 22404630 Free PMC article. Review.
-
Breast cancer genes PSMC3IP and EPSTI1 play a role in apoptosis regulation.PLoS One. 2015 Jan 15;10(1):e0115352. doi: 10.1371/journal.pone.0115352. eCollection 2015. PLoS One. 2015. PMID: 25590583 Free PMC article.
-
GT198 Splice Variants Display Dominant-Negative Activities and Are Induced by Inactivating Mutations.Genes Cancer. 2013 Jan;4(1-2):26-38. doi: 10.1177/1947601913486345. Genes Cancer. 2013. PMID: 23946869 Free PMC article.
-
MicroRNAs affecting the susceptibility of melanoma cells to CD8+ T cell-mediated cytolysis.Clin Transl Med. 2023 Feb;13(2):e1186. doi: 10.1002/ctm2.1186. Clin Transl Med. 2023. PMID: 36718025 Free PMC article.
-
Oncoprotein GT198 vaccination delays tumor growth in MMTV-PyMT mice.Cancer Lett. 2020 Apr 28;476:57-66. doi: 10.1016/j.canlet.2020.02.005. Epub 2020 Feb 12. Cancer Lett. 2020. PMID: 32061755 Free PMC article.
References
-
- Holdcraft RW, Braun RE 2004 Hormonal regulation of spermatogenesis. Int J Androl 27:335–342 - PubMed
-
- Dehm SM, Tindall DJ 2006 Molecular regulation of androgen action in prostate cancer. J Cell Biochem 99:333–344 - PubMed
-
- Prescott J, Coetzee GA 2006 Molecular chaperones throughout the life cycle of the androgen receptor. Cancer Lett 231:12–19 - PubMed
-
- Culig Z, Comuzzi B, Steiner H, Bartsch G, Hobisch A 2004 Expression and function of androgen receptor coactivators in prostate cancer. J Steroid Biochem Mol Biol 92:265–271 - PubMed
-
- Ikonen T, Palvimo JJ, Jänne OA 1997 Interaction between the amino- and carboxyl-terminal regions of the rat androgen receptor modulates transcriptional activity and is influenced by nuclear receptor coactivators. J Biol Chem 272:29821–29828 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
