

The type III transforming growth factor-beta receptor negatively regulates nuclear factor kappa B signaling through its interaction with beta-arrestin2
- PMID: 19325136
- PMCID: PMC2718069
- DOI: 10.1093/carcin/bgp071
The type III transforming growth factor-beta receptor negatively regulates nuclear factor kappa B signaling through its interaction with beta-arrestin2
Abstract
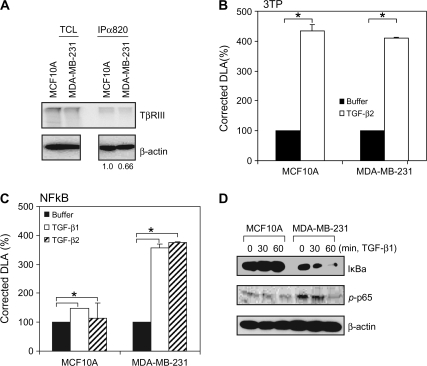
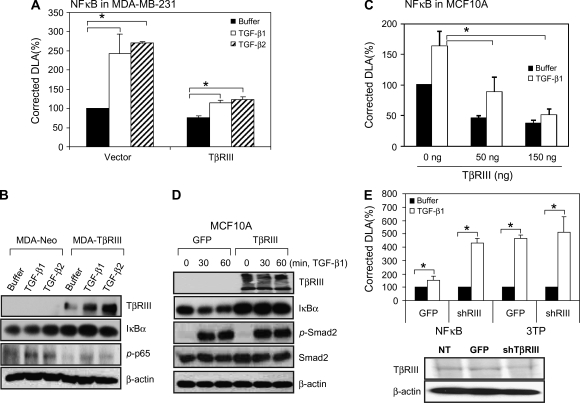
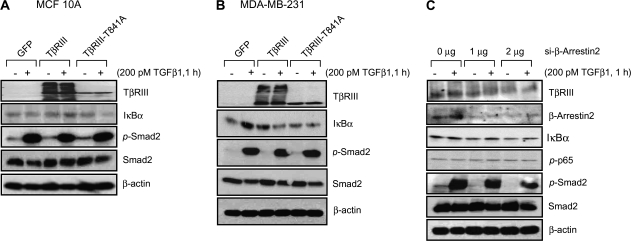
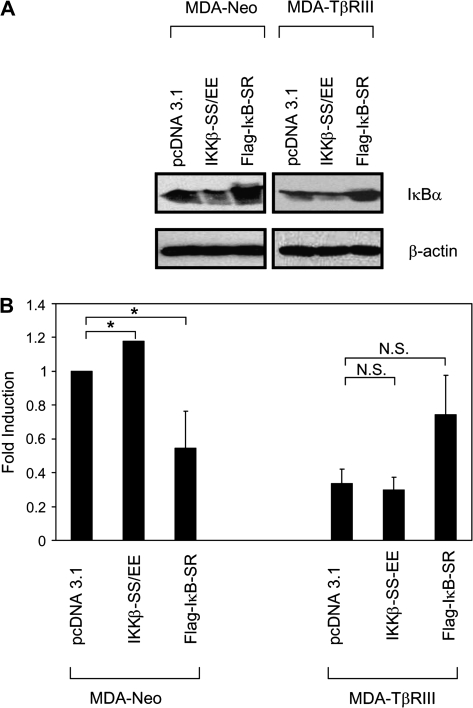
Transforming growth factor-beta (TGF-beta) increases or decreases nuclear factor kappa B (NFkappaB) signaling in a context-dependent manner through mechanisms that remain to be defined. The type III transforming growth factor-beta receptor (TbetaRIII) is a TGF-beta superfamily co-receptor with emerging roles in both mediating and regulating TGF-beta superfamily signaling. We have previously reported a novel interaction of TbetaRIII with the scaffolding protein, beta-arrestin2, which results in TbetaRIII internalization and downregulation of TGF-beta signaling. beta-arrestin2 also scaffolds interacting receptors with the mitogen-activated protein kinase and NFkappaB-signaling pathways. Here, we demonstrate that TbetaRIII, through its interaction with beta-arrestin2, negatively regulates NFkappaB signaling in MCF10A breast epithelial and MDA-MB-231 breast cancer cells. Increasing TbetaRIII expression reduced NFkappaB-mediated transcriptional activation and IkappaBalpha degradation, whereas a TbetaRIII mutant unable to interact with beta-arrestin2, TbetaRIII-T841A, had no effect. In a reciprocal manner, short hairpin RNA-mediated silencing of either TbetaRIII expression or beta-arrestin2 expression increased NFkappaB-mediated transcriptional activation and IkappaBalpha degradation. Functionally, TbetaRIII-mediated repression of NFkappaB signaling is important for TbetaRIII-mediated inhibition of breast cancer cell migration. These studies define a mechanism through which TbetaRIII regulates NFkappaB signaling and expand the roles of this TGF-beta superfamily co-receptor in regulating epithelial cell homeostasis.
Figures
References
-
- Bierie B, et al. Tumour microenvironment: TGFbeta: the molecular Jekyll and Hyde of cancer. Nat. Rev. Cancer. 2006;6:506–520. - PubMed
-
- Elliott RL, et al. Role of transforming growth factor Beta in human cancer. J. Clin. Oncol. 2005;23:2078–2093. - PubMed
-
- Gordon KJ, et al. Role of transforming growth factor-beta superfamily signaling pathways in human disease. Biochim. Biophys. Acta. 2008;1782:197–228. - PubMed
-
- Kirkbride KC, et al. Cell-surface co-receptors: emerging roles in signaling and human disease. Trends Biochem. Sci. 2005;30:611–621. - PubMed
-
- Massague J, et al. TGFbeta signaling in growth control, cancer, and heritable disorders. Cell. 2000;103:295–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
