

Neurophysiological bases of exponential sensory decay and top-down memory retrieval: a model
- PMID: 19325713
- PMCID: PMC2659975
- DOI: 10.3389/neuro.10.004.2009
Neurophysiological bases of exponential sensory decay and top-down memory retrieval: a model
Abstract
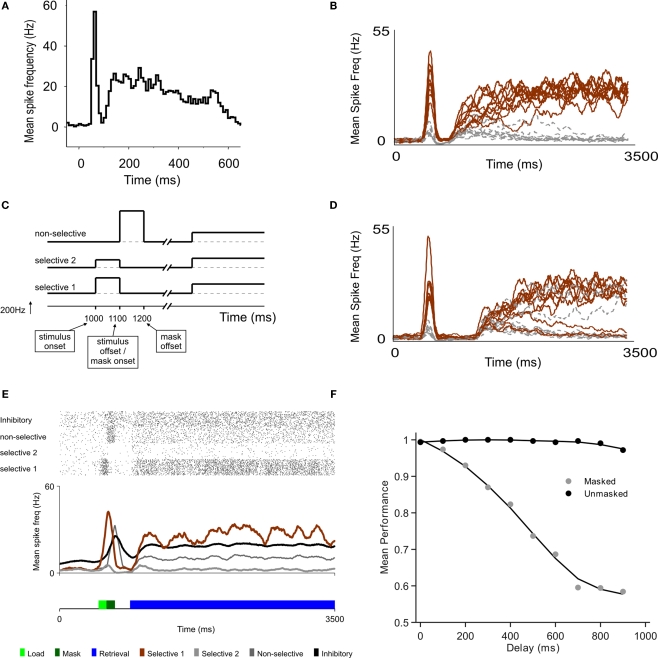
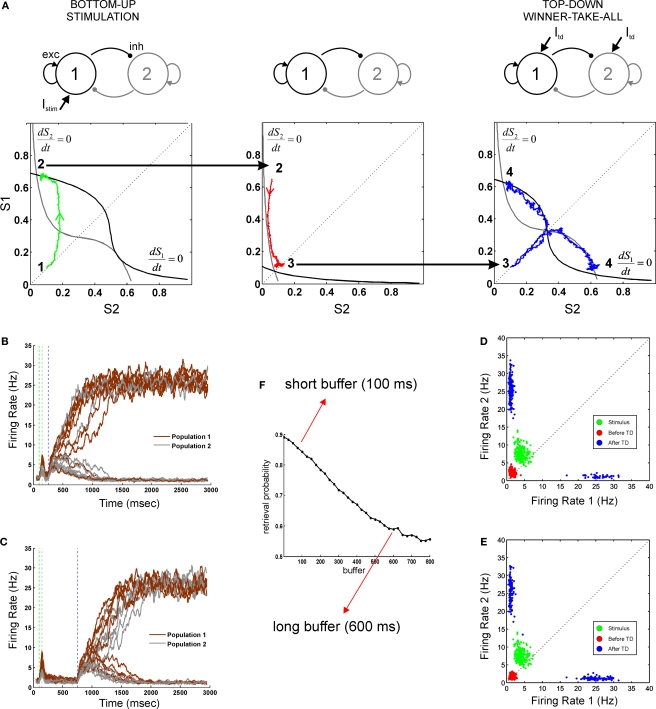
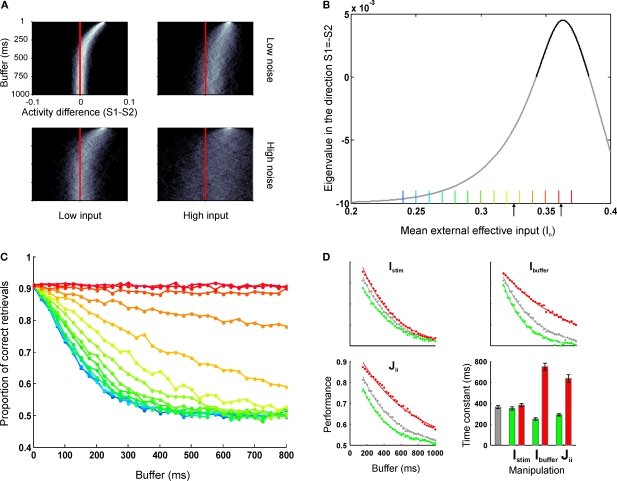
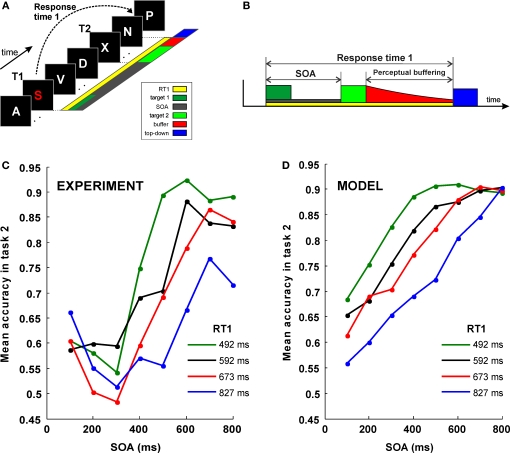
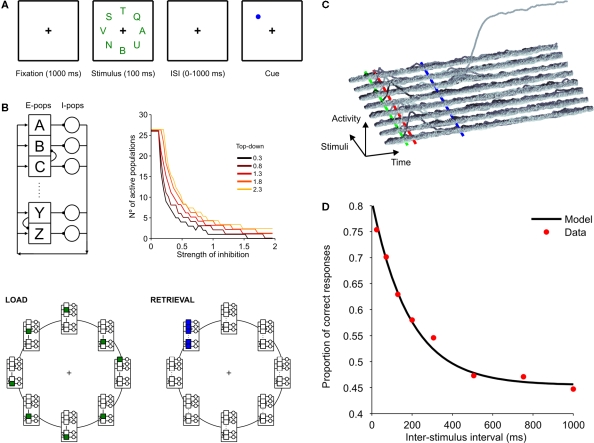
Behavioral observations suggest that multiple sensory elements can be maintained for a short time, forming a perceptual buffer which fades after a few hundred milliseconds. Only a subset of this perceptual buffer can be accessed under top-down control and broadcasted to working memory and consciousness. In turn, single-cell studies in awake-behaving monkeys have identified two distinct waves of response to a sensory stimulus: a first transient response largely determined by stimulus properties and a second wave dependent on behavioral relevance, context and learning. Here we propose a simple biophysical scheme which bridges these observations and establishes concrete predictions for neurophsyiological experiments in which the temporal interval between stimulus presentation and top-down allocation is controlled experimentally. Inspired in single-cell observations, the model involves a first transient response and a second stage of amplification and retrieval, which are implemented biophysically by distinct operational modes of the same circuit, regulated by external currents. We explicitly investigated the neuronal dynamics, the memory trace of a presented stimulus and the probability of correct retrieval, when these two stages were bracketed by a temporal gap. The model predicts correctly the dependence of performance with response times in interference experiments suggesting that sensory buffering does not require a specific dedicated mechanism and establishing a direct link between biophysical manipulations and behavioral observations leading to concrete predictions.
Keywords: attentional blink; attractor networks; dual-task interference; iconic memory; stochastic processes.
Figures
Similar articles
-
Beyond a mask and against the bottleneck: retroactive dual-task interference during working memory consolidation of a masked visual target.J Exp Psychol Gen. 2014 Jun;143(3):1409-1427. doi: 10.1037/a0035257. Epub 2013 Dec 23. J Exp Psychol Gen. 2014. PMID: 24364683
-
Hydrocortisone accelerates the decay of iconic memory traces: on the modulation of executive and stimulus-driven constituents of sensory information maintenance.Psychoneuroendocrinology. 2015 Mar;53:148-58. doi: 10.1016/j.psyneuen.2014.12.016. Epub 2015 Jan 3. Psychoneuroendocrinology. 2015. PMID: 25618593
-
Bottom-Up and Top-Down Factors Differentially Influence Stimulus Representations Across Large-Scale Attentional Networks.J Neurosci. 2018 Mar 7;38(10):2495-2504. doi: 10.1523/JNEUROSCI.2724-17.2018. Epub 2018 Feb 2. J Neurosci. 2018. PMID: 29437930 Free PMC article.
-
Fuzzy-trace theory: dual processes in memory, reasoning, and cognitive neuroscience.Adv Child Dev Behav. 2001;28:41-100. doi: 10.1016/s0065-2407(02)80062-3. Adv Child Dev Behav. 2001. PMID: 11605365 Review.
-
The offline stream of conscious representations.Philos Trans R Soc Lond B Biol Sci. 2018 Sep 19;373(1755):20170349. doi: 10.1098/rstb.2017.0349. Philos Trans R Soc Lond B Biol Sci. 2018. PMID: 30061463 Free PMC article. Review.
Cited by
-
Will the real resource theory please stand up! Vigilance is a renewable resource and should be modeled as such.Exp Brain Res. 2023 May;241(5):1263-1270. doi: 10.1007/s00221-023-06604-x. Epub 2023 Mar 30. Exp Brain Res. 2023. PMID: 36997721
-
A temporal bottleneck in the language comprehension network.J Neurosci. 2012 Jun 27;32(26):9089-102. doi: 10.1523/JNEUROSCI.5685-11.2012. J Neurosci. 2012. PMID: 22745508 Free PMC article.
-
It's hard to forget: resetting memory in delay-match-to-multiple-image tasks.Front Hum Neurosci. 2013 Nov 14;7:765. doi: 10.3389/fnhum.2013.00765. eCollection 2013. Front Hum Neurosci. 2013. PMID: 24294199 Free PMC article.
-
Top-down influences of written text on perceived clarity of degraded speech.J Exp Psychol Hum Percept Perform. 2014 Feb;40(1):186-99. doi: 10.1037/a0033206. Epub 2013 Jun 10. J Exp Psychol Hum Percept Perform. 2014. PMID: 23750966 Free PMC article. Clinical Trial.
-
Relation of frontal N100 to psychopathy-related differences in selective attention.Biol Psychol. 2014 Dec;103:107-16. doi: 10.1016/j.biopsycho.2014.08.012. Epub 2014 Aug 29. Biol Psychol. 2014. PMID: 25179538 Free PMC article.
References
-
- Averbach E., Coriell A. S. (1961). Short-term memory in vision. Bell Syst. Tech. J. 40, 309–328
-
- Baars B. J. (1989). A Cognitive Theory of Consciousness. Cambridge, Cambridge University Press
LinkOut - more resources
Full Text Sources
Miscellaneous