

Genomic evidence for the evolution of Streptococcus equi: host restriction, increased virulence, and genetic exchange with human pathogens
- PMID: 19325880
- PMCID: PMC2654543
- DOI: 10.1371/journal.ppat.1000346
Genomic evidence for the evolution of Streptococcus equi: host restriction, increased virulence, and genetic exchange with human pathogens
Abstract
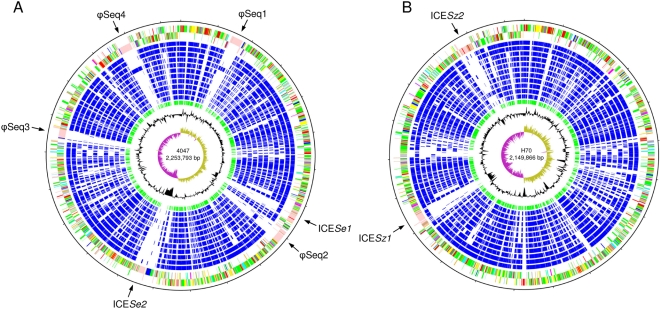
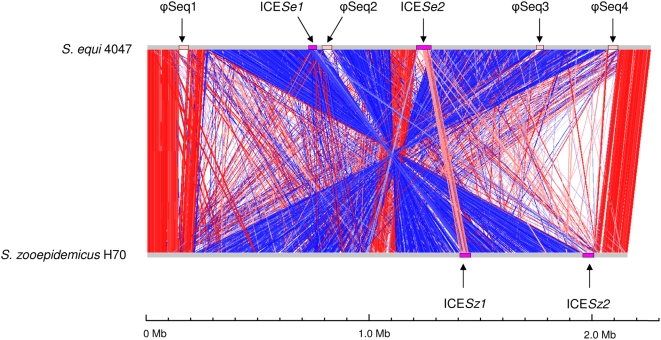
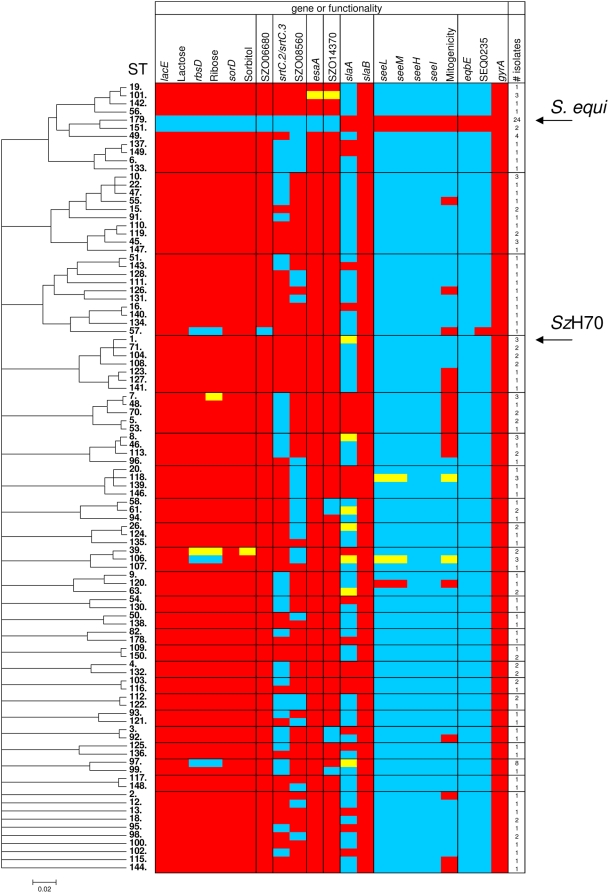
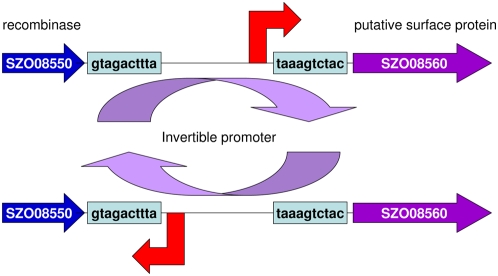
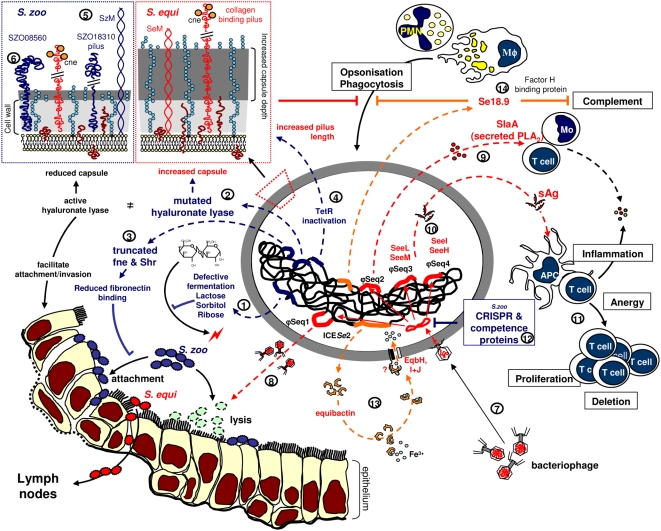
The continued evolution of bacterial pathogens has major implications for both human and animal disease, but the exchange of genetic material between host-restricted pathogens is rarely considered. Streptococcus equi subspecies equi (S. equi) is a host-restricted pathogen of horses that has evolved from the zoonotic pathogen Streptococcus equi subspecies zooepidemicus (S. zooepidemicus). These pathogens share approximately 80% genome sequence identity with the important human pathogen Streptococcus pyogenes. We sequenced and compared the genomes of S. equi 4047 and S. zooepidemicus H70 and screened S. equi and S. zooepidemicus strains from around the world to uncover evidence of the genetic events that have shaped the evolution of the S. equi genome and led to its emergence as a host-restricted pathogen. Our analysis provides evidence of functional loss due to mutation and deletion, coupled with pathogenic specialization through the acquisition of bacteriophage encoding a phospholipase A(2) toxin, and four superantigens, and an integrative conjugative element carrying a novel iron acquisition system with similarity to the high pathogenicity island of Yersinia pestis. We also highlight that S. equi, S. zooepidemicus, and S. pyogenes share a common phage pool that enhances cross-species pathogen evolution. We conclude that the complex interplay of functional loss, pathogenic specialization, and genetic exchange between S. equi, S. zooepidemicus, and S. pyogenes continues to influence the evolution of these important streptococci.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Jorm LR, Love DN, Bailey GD, McKay GM, Briscoe DA. Genetic structure of populations of beta-haemolytic Lancefield group C streptococci from horses and their association with disease. Res Vet Sci. 1994;57:292–299. - PubMed
-
- Webb K, Jolley KA, Mitchell Z, Robinson C, Newton JR, et al. Development of an unambiguous and discriminatory multilocus sequence typing scheme for the Streptococcus zooepidemicus group. Microbiology. 2008;154:3016–3024. - PubMed
-
- Alber J, El-Sayed A, Estoepangestie S, Lammler C, Zschock M. Dissemination of the superantigen encoding genes seeL, seeM, szeL and szeM in Streptococcus equi subsp. equi and Streptococcus equi subsp. zooepidemicus. Vet Microbiol. 2005;109:135–141. - PubMed
-
- Artiushin SC, Timoney JF, Sheoran AS, Muthupalani SK. Characterization and immunogenicity of pyrogenic mitogens SePE-H and SePE-I of Streptococcus equi. Microb Pathog. 2002;32:71–85. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
