

Not just for bimodal neurons anymore: the contribution of unimodal neurons to cortical multisensory processing
- PMID: 19326204
- PMCID: PMC2854489
- DOI: 10.1007/s10548-009-0088-3
Not just for bimodal neurons anymore: the contribution of unimodal neurons to cortical multisensory processing
Abstract
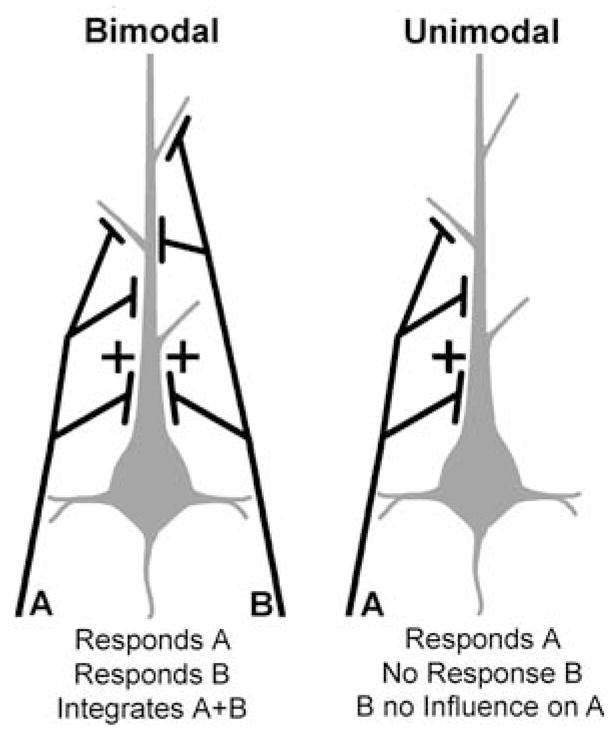
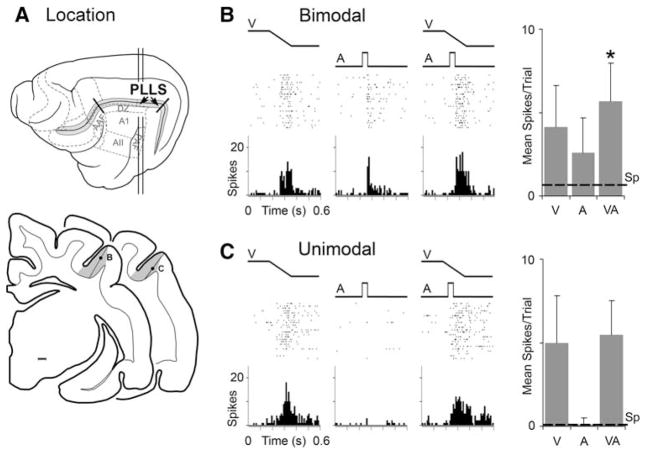
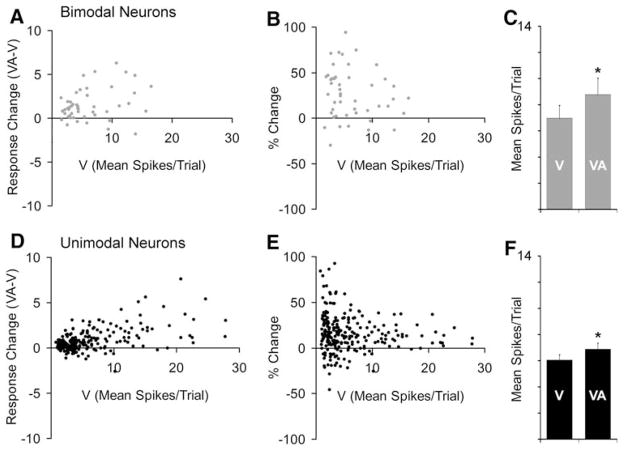
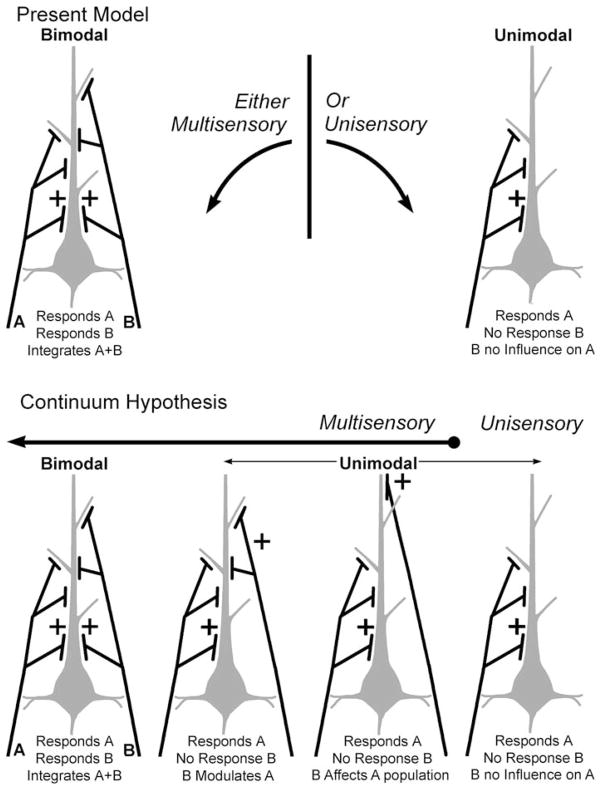
Traditionally, neuronal studies of multisensory processing proceeded by first identifying neurons that were overtly multisensory (e.g., bimodal, trimodal) and then testing them. In contrast, the present study examined, without precondition, neurons in an extrastriate visual area of the cat for their responses to separate (visual, auditory) and combined-modality (visual and auditory) stimulation. As expected, traditional bimodal forms of multisensory neurons were identified. In addition, however, many neurons that were activated only by visual stimulation (i.e., unimodal) had that response modulated by the presence of an auditory stimulus. Some unimodal neurons showed multisensory responses that were statistically different from their visual response. Other unimodal neurons had subtle multisensory effects that were detectable only at the population level. Most surprisingly, these non-bimodal neurons generated more than twice the multisensory signal in the PLLS than did the bimodal neurons. These results expand the range of multisensory convergence patterns beyond that of the bimodal neuron. However, rather than characterize a separate class of multisensory neurons, unimodal multisensory neurons may actually represent an intermediary form of multisensory convergence that exists along the functional continuum between unisensory neurons, at one end, and fully bimodal neurons at the other.
Figures

References
-
- Allman BL, Meredith MA. Multisensory processing in ‘unimodal’ neurons: cross-modal subthreshold auditory effects in cat extrastriate visual cortex. J Neurophysiol. 2007;98:545–549. - PubMed
-
- Barraclough NE, Xiao D, Baker CI, Oram MW, Perrett DI. Integration of visual and auditory information by Superior Temporal Sulcus neurons responsive to the sight of actions. J Cognitive Neurosci. 2005;17:377–391. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
