

Anchorage-independent growth of breast carcinoma cells is mediated by serum exosomes
- PMID: 19327352
- PMCID: PMC2742412
- DOI: 10.1016/j.yexcr.2009.03.010
Anchorage-independent growth of breast carcinoma cells is mediated by serum exosomes
Abstract
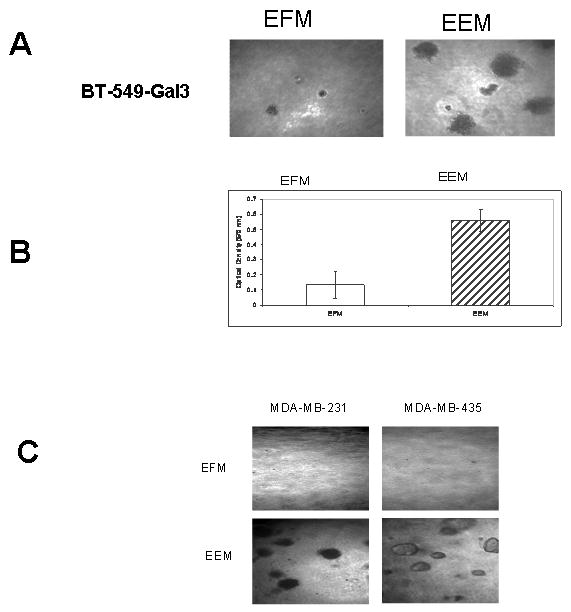
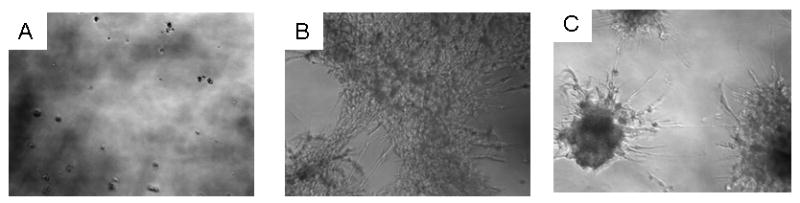
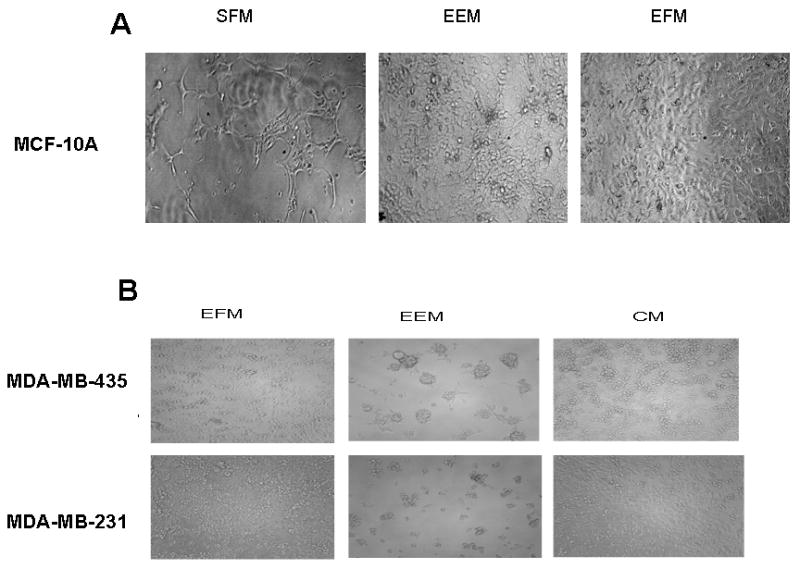
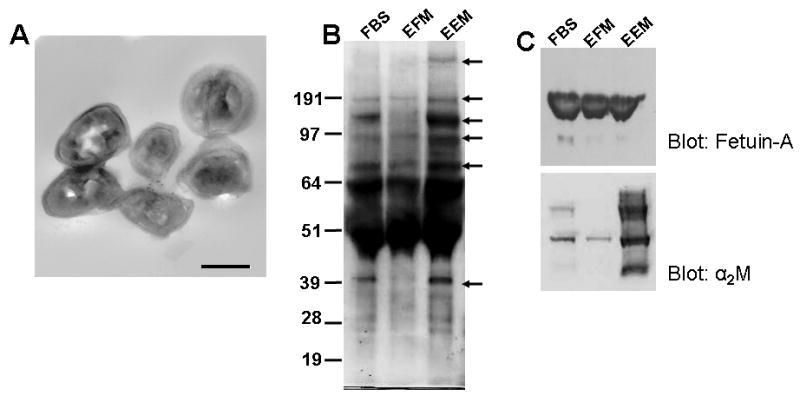
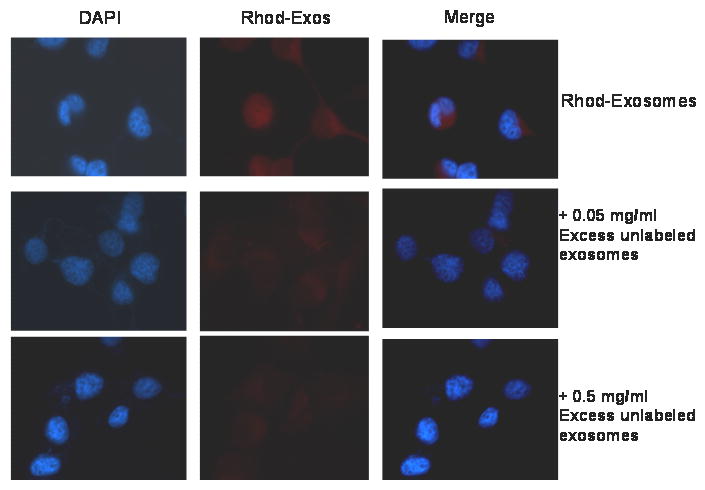
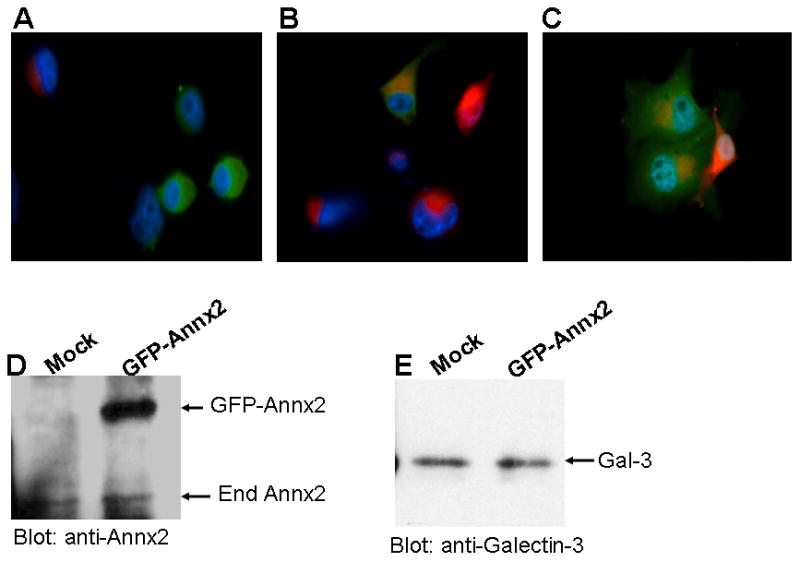
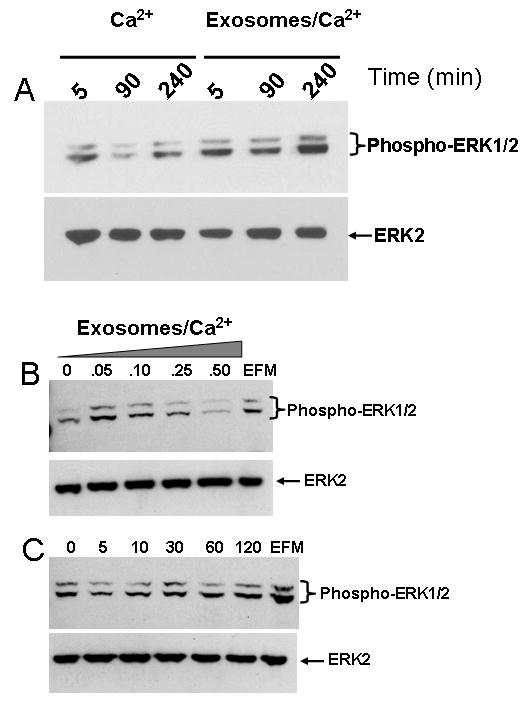
We hereby report studies that suggest a role for serum exosomes in the anchorage-independent growth (AIG) of tumor cells. In AIG assays, fetal bovine serum is one of the critical ingredients. We therefore purified exosomes from fetal bovine serum and examined their potential to promote growth of breast carcinoma cells in soft agar and Matrigel after reconstituting them into growth medium (EEM). In all the assays, viable colonies were formed only in the presence of exosomes. Some of the exosomal proteins we identified, have been documented by others and could be considered exosomal markers. Labeled purified exosomes were up-taken by the tumor cells, a process that could be competed out with excess unlabeled vesicles. Our data also suggested that once endocytosed by a cell, the exosomes could be recycled back to the conditioned medium from where they can be up-taken by other cells. We also demonstrated that low concentrations of exosomes activate MAP kinases, suggesting a mechanism by which they maintain the growth of the tumor cells in soft agar. Taken together, our data demonstrate that serum exosomes form a growth promoting platform for AIG of tumor cells and may open a new vista into cancer cell growth in vivo.
Figures
References
-
- Klein EA, Yung Y, Castagnino P, Kothapalli D, Assoian RK. Cell adhesion, cellular tension, and cell cycle control. Methods Enzymol. 2007;426:155–175. - PubMed
-
- Reichelt J. Mechanotransduction of keratinocytes in culture and in the epidermis. Eur J Cell Biol. 2007;86:807–816. - PubMed
-
- Freedman VH, Shin SI. Cellular tumorigenicity in nude mice: correlation with cell growth in semi-solid medium. Cell. 1974;3:355–359. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
