

Live-cell imaging demonstrates extracellular matrix degradation in association with active cathepsin B in caveolae of endothelial cells during tube formation
- PMID: 19331819
- PMCID: PMC2677760
- DOI: 10.1016/j.yexcr.2009.01.021
Live-cell imaging demonstrates extracellular matrix degradation in association with active cathepsin B in caveolae of endothelial cells during tube formation
Abstract
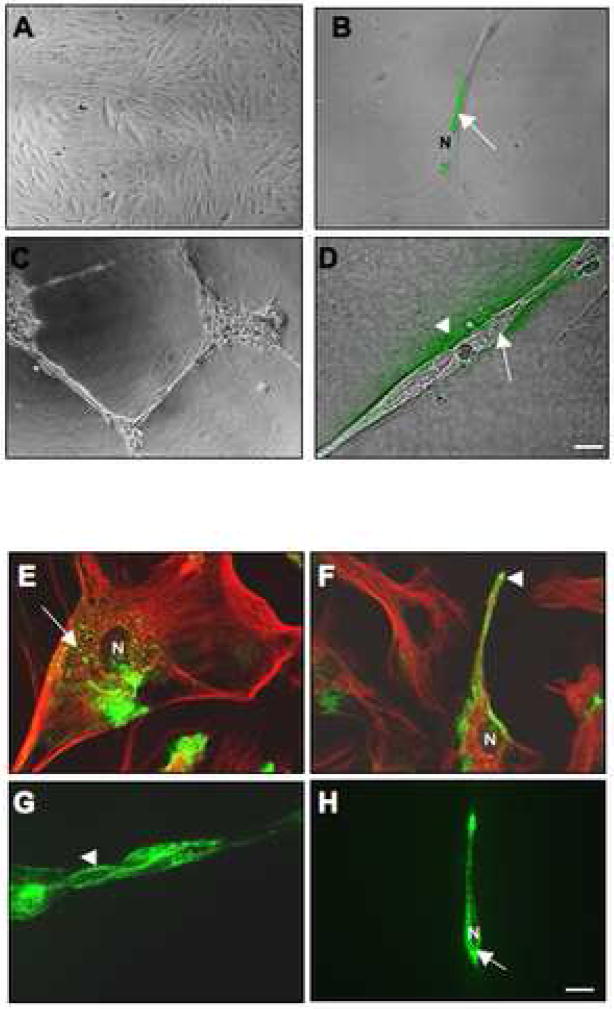
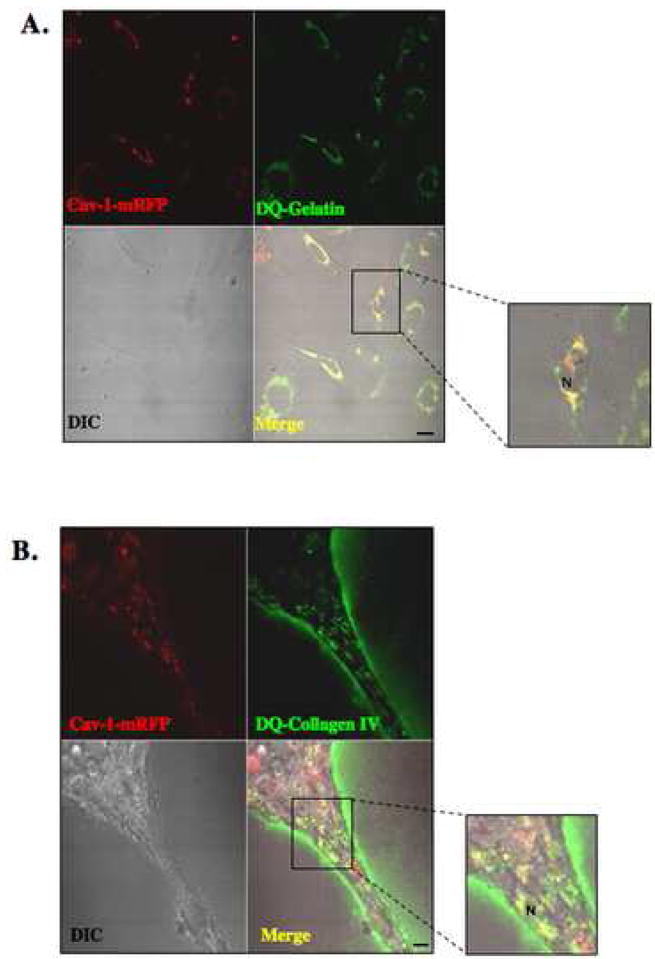
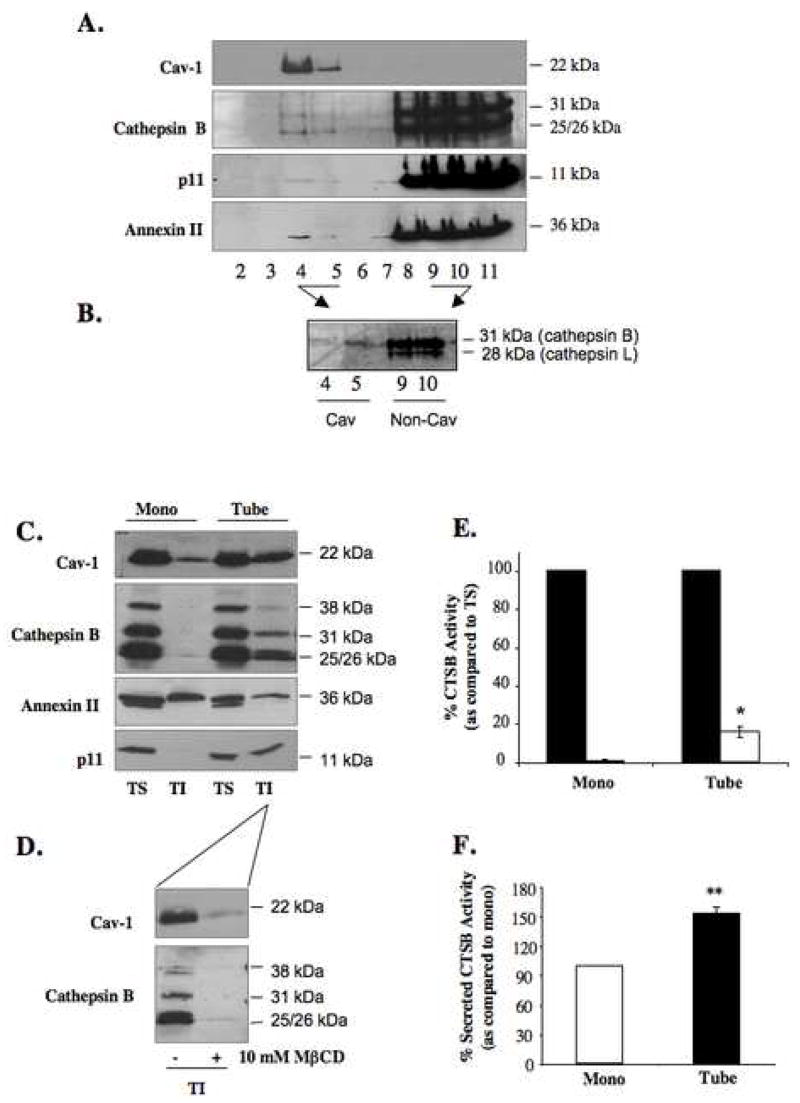
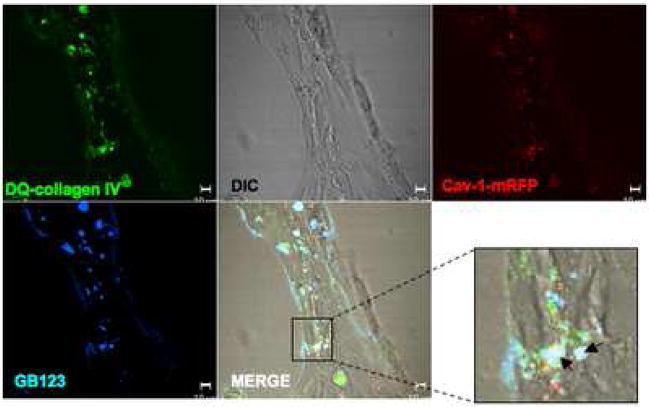
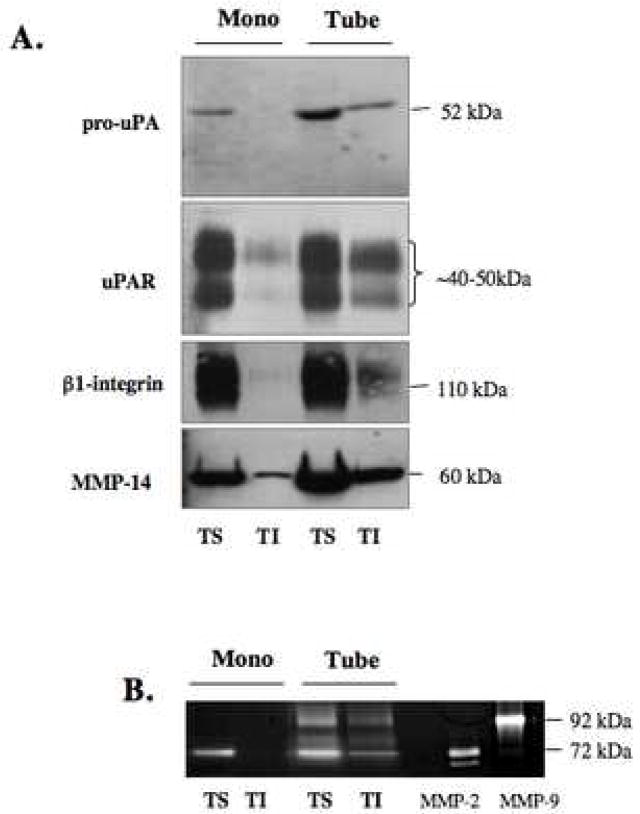
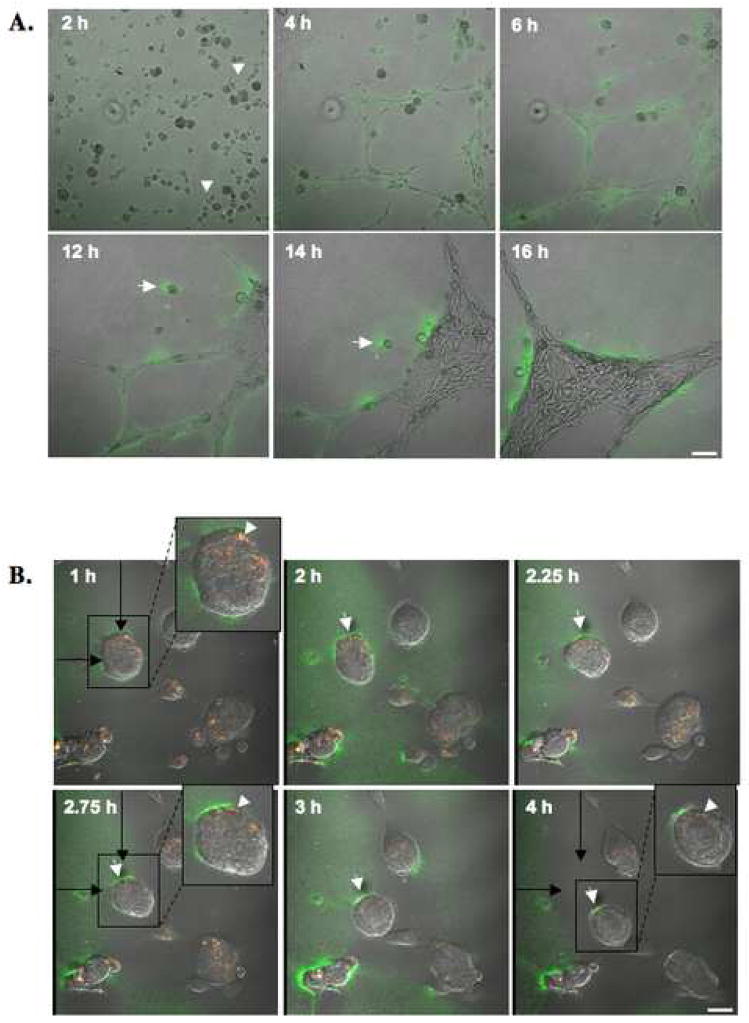
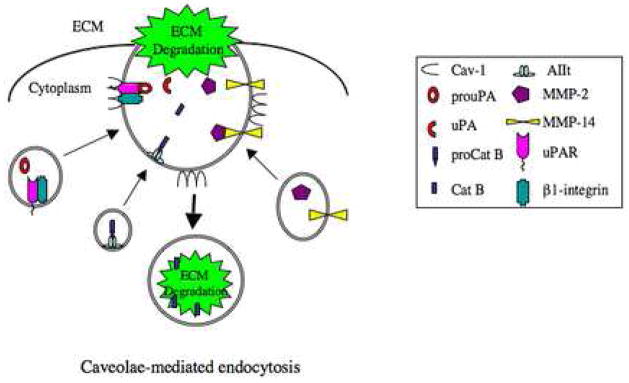
Localization of proteases to the surface of endothelial cells and remodeling of the extracellular matrix (ECM) are essential to endothelial cell tube formation and angiogenesis. Here, we partially localized active cathepsin B and its cell surface binding partners, S100A/p11 (p11) of the annexin II heterotetramer (AIIt), to caveolae of human umbilical vein endothelial cells (HUVEC). Via a live-cell proteolysis assay, we observed that degradation products of quenched-fluorescent (DQ)-proteins (i.e. gelatin and collagen IV) colocalized intracellularly with caveolin-1 (cav-1) of HUVEC grown in either monolayer cultures or in vitro tube formation assays. Activity-based probes that bind covalently to active cysteine cathepsins and degradation products of DQ-collagen IV partially localized to intracellular vesicles that contained cav-1 and active cysteine cathepsins. Biochemical analyses revealed that the distribution of active cathepsin B in caveolar fractions increased during in vitro tube formation. Pro-uPA, uPAR, MMP-2 and MMP-14, which have been linked with cathepsin B to ECM degradation pathways, were also found to increase in caveolar fractions during in vitro tube formation. Our findings are the first to demonstrate through live-cell imaging ECM degradation in association with active cathepsin B in caveolae of endothelial cells during tube formation.
Figures
References
-
- Risau W. Mechanisms of angiogenesis. Nature. 1997;386:671–674. - PubMed
-
- van Hinsbergh VW, Engelse MA, Quax PH. Pericellular proteases in angiogenesis and vasculogenesis. Arterioscler Thromb Vasc Biol. 2006;26:716–728. - PubMed
-
- Pepper MS. Role of the matrix metalloproteinase and plasminogen activator-plasmin systems in angiogenesis. Arterioscler Thromb Vasc Biol. 2001;21:1104–1117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
