

Gating of the CFTR Cl- channel by ATP-driven nucleotide-binding domain dimerisation
- PMID: 19332488
- PMCID: PMC2697289
- DOI: 10.1113/jphysiol.2009.171595
Gating of the CFTR Cl- channel by ATP-driven nucleotide-binding domain dimerisation
Abstract
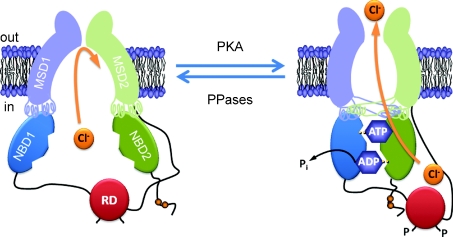
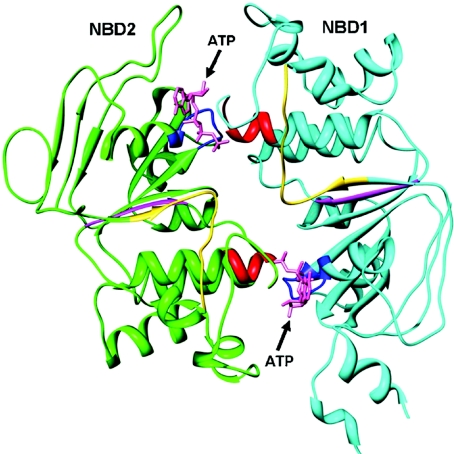
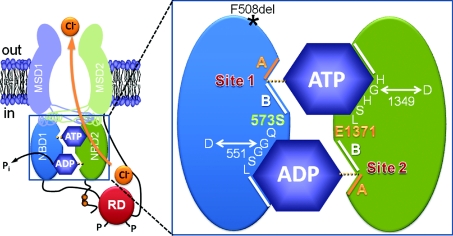
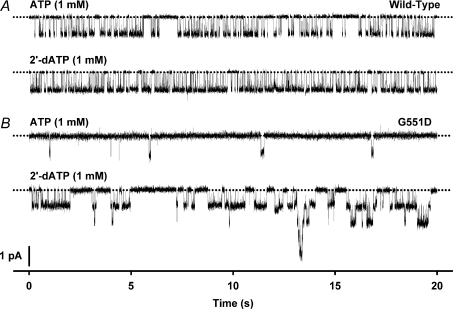
The cystic fibrosis transmembrane conductance regulator (CFTR) plays a fundamental role in fluid and electrolyte transport across epithelial tissues. Based on its structure, function and regulation, CFTR is an ATP-binding cassette (ABC) transporter. These transporters are assembled from two membrane-spanning domains (MSDs) and two nucleotide-binding domains (NBDs). In the vast majority of ABC transporters, the NBDs form a common engine that utilises the energy of ATP hydrolysis to pump a wide spectrum of substrates through diverse transmembrane pathways formed by the MSDs. By contrast, in CFTR the MSDs form a pathway for passive anion flow that is gated by cycles of ATP binding and hydrolysis by the NBDs. Here, we consider how the interaction of ATP with two ATP-binding sites, formed by the NBDs, powers conformational changes in CFTR structure to gate the channel pore. We explore how conserved sequences from both NBDs form ATP-binding sites at the interface of an NBD dimer and highlight the distinct roles that each binding site plays during the gating cycle. Knowledge of how ATP gates the CFTR Cl- channel is critical for understanding CFTR's physiological role, its malfunction in disease and the mechanism of action of small molecules that modulate CFTR channel gating.
Figures
References
-
- Aleksandrov AA, Aleksandrov L, Riordan JR. Nucleoside triphosphate pentose ring impact on CFTR gating and hydrolysis. FEBS Lett. 2002;518:183–188. - PubMed
-
- Aleksandrov AA, Aleksandrov LA, Riordan JR. CFTR (ABCC7) is a hydrolyzable-ligand-gated channel. Pflügers Arch. 2007;453:693–702. - PubMed
-
- Aleksandrov L, Aleksandrov A, Riordan JR. Mg2+-dependent ATP occlusion at the first nucleotide-binding domain (NBD1) of CFTR does not require the second (NBD2) Biochem J. 2008;416:129–136. - PubMed
-
- Anderson MP, Berger HA, Rich DP, Gregory RJ, Smith AE, Welsh MJ. Nucleoside triphosphates are required to open the CFTR chloride channel. Cell. 1991;67:775–784. - PubMed
-
- Anderson MP, Welsh MJ. Regulation by ATP and ADP of CFTR chloride channels that contain mutant nucleotide-binding domains. Science. 1992;257:1701–1704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources