

Strabismus regulates asymmetric cell divisions and cell fate determination in the mouse brain
- PMID: 19332887
- PMCID: PMC2700512
- DOI: 10.1083/jcb.200807073
Strabismus regulates asymmetric cell divisions and cell fate determination in the mouse brain
Abstract
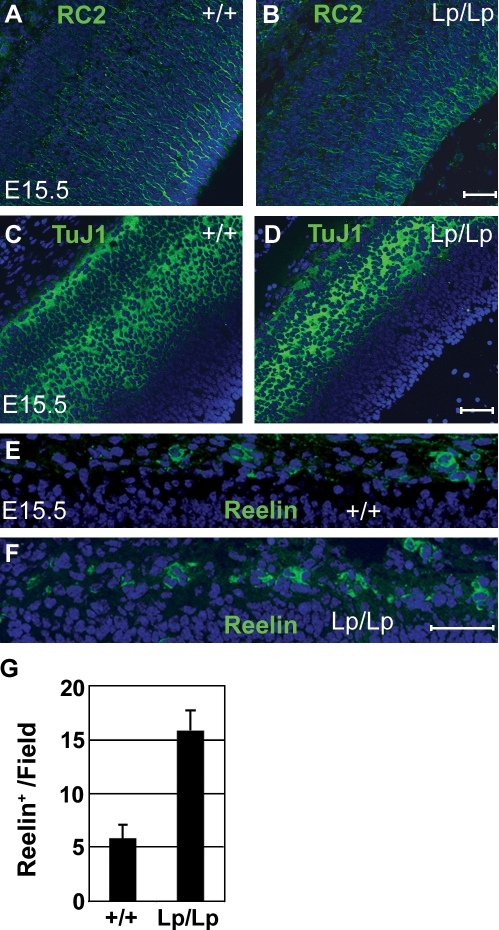
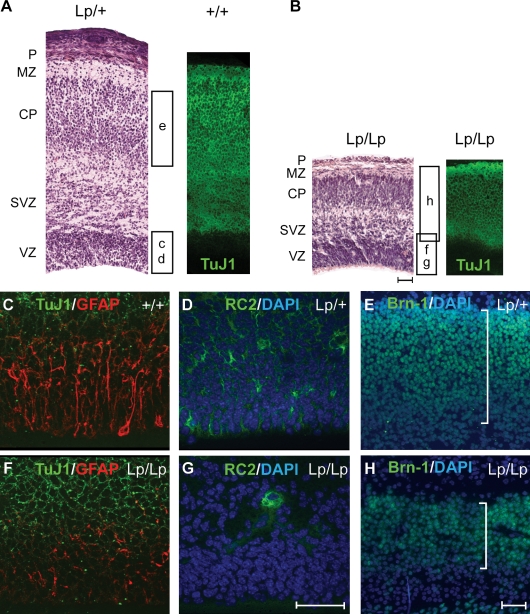
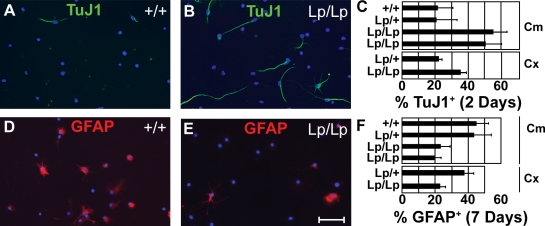
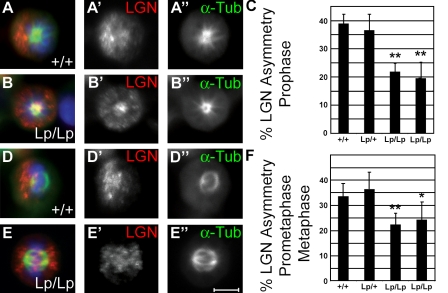
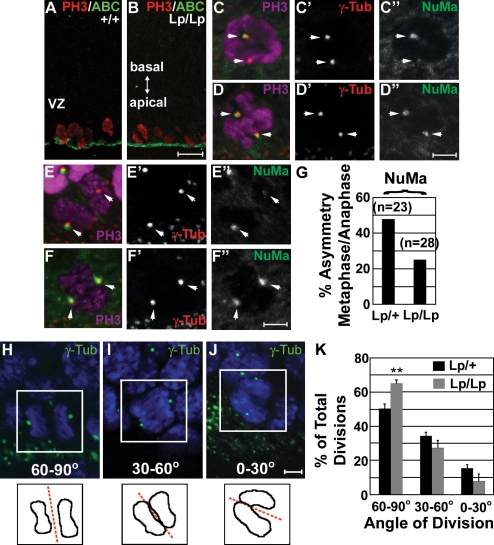
The planar cell polarity (PCP) pathway organizes the cytoskeleton and polarizes cells within embryonic tissue. We investigate the relationship between PCP signaling and cell fate determination during asymmetric division of neural progenitors (NPs) in mouse embryos. The cortex of Lp/Lp (Loop-tail) mice deficient in the essential PCP mediator Vangl2, homologue of Drosophila melanogaster Strabismus (Stbm), revealed precocious differentiation of neural progenitors into early-born neurons at the expense of late-born neurons and glia. Although Lp/Lp NPs were easily maintained in vitro, they showed premature differentiation and loss of asymmetric distribution of Leu-Gly-Asn-enriched protein (LGN)/partner of inscuteable (Pins), a regulator of mitotic spindle orientation. Furthermore, we observed a decreased frequency in asymmetric distribution of the LGN target nuclear mitotic apparatus protein (NuMa) in Lp/Lp cortical progenitors in vivo. This was accompanied by an increase in the number of vertical cleavage planes typically associated with equal daughter cell identities. These findings suggest that Stbm/Vangl2 functions to maintain cortical progenitors and regulates mitotic spindle orientation during asymmetric divisions in the vertebrate brain.
Figures
References
-
- Bellaiche Y., Beaudoin-Massiani O., Stuttem I., Schweisguth F. 2004. The planar cell polarity protein Strabismus promotes Pins anterior localization during asymmetric division of sensory organ precursor cells in Drosophila.Development. 131:469–478 - PubMed
-
- Betschinger J., Knoblich J.A. 2004. Dare to be different: asymmetric cell division in Drosophila, C. elegans and vertebrates.Curr. Biol. 14:R674–R685 - PubMed
-
- Blumer J.B., Chandler L.J., Lanier S.M. 2002. Expression analysis and subcellular distribution of the two G-protein regulators AGS3 and LGN indicate distinct functionality. Localization of LGN to the midbody during cytokinesis.J. Biol. Chem. 277:15897–15903 - PubMed
-
- Bond J., Roberts E., Mochida G.H., Hampshire D.J., Scott S., Askham J.M., Springell K., Mahadevan M., Crow Y.J., Markham A.F., et al. 2002. ASPM is a major determinant of cerebral cortical size.Nat. Genet. 32:316–320 - PubMed
-
- Brott B.K., Sokol S.Y. 2005. A vertebrate homolog of the cell cycle regulator Dbf4 is an inhibitor of Wnt signaling required for heart development.Dev. Cell. 8:703–715 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
