

Estrogen and progesterone receptors: from molecular structures to clinical targets
- PMID: 19333551
- PMCID: PMC11115849
- DOI: 10.1007/s00018-009-0017-3
Estrogen and progesterone receptors: from molecular structures to clinical targets
Abstract
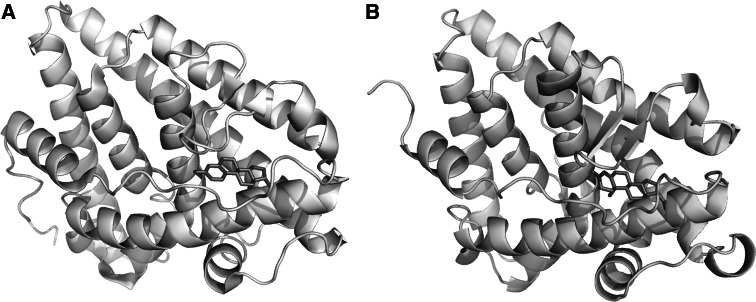
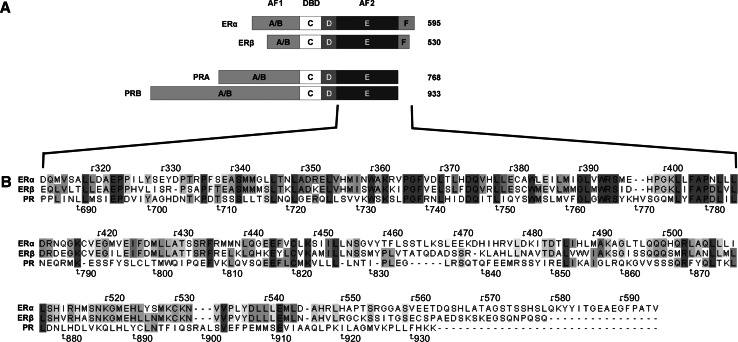
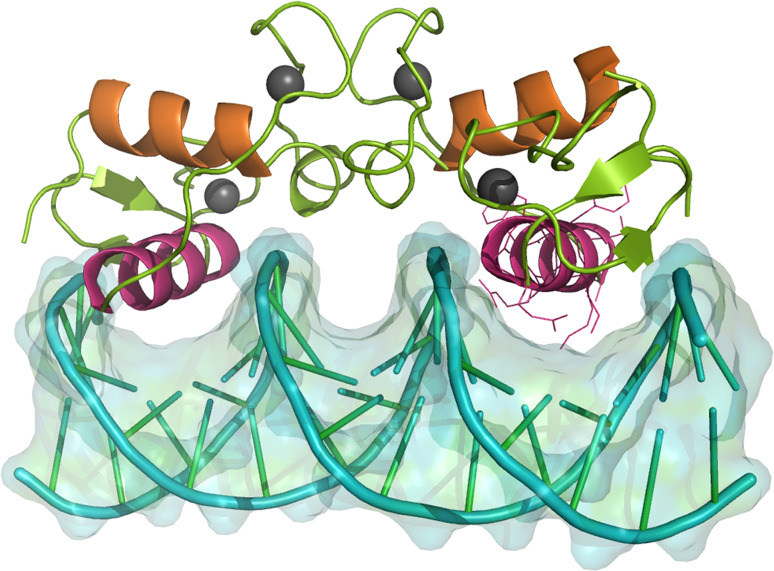
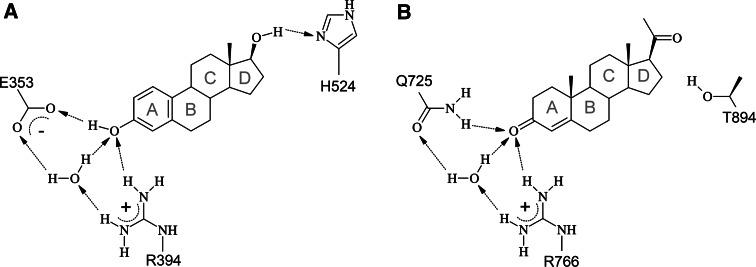
Research involving estrogen and progesterone receptors (ER and PR) have greatly contributed to our understanding of cell signaling and transcriptional regulation. In addition to the classical ER and PR nuclear actions, new signaling pathways have recently been identified due to ER and PR association with cell membranes and signal transduction proteins. Bio-informatics has unveiled how ER and PR recognize their ligands, selective modulators and co-factors, which has helped to implement them as key targets in the treatment of benign and malignant tumors. Knowledge regarding ER and PR is vast and complex; therefore, this review will focus on their isoforms, signaling pathways, co-activators and co-repressors, which lead to target gene regulation. Moreover it will highlight ER and PR involvement in benign and malignant diseases as well as pharmacological substances influencing cell signaling and provide established and new structural insights into the mechanism of activation and inhibition of these receptors.
Figures


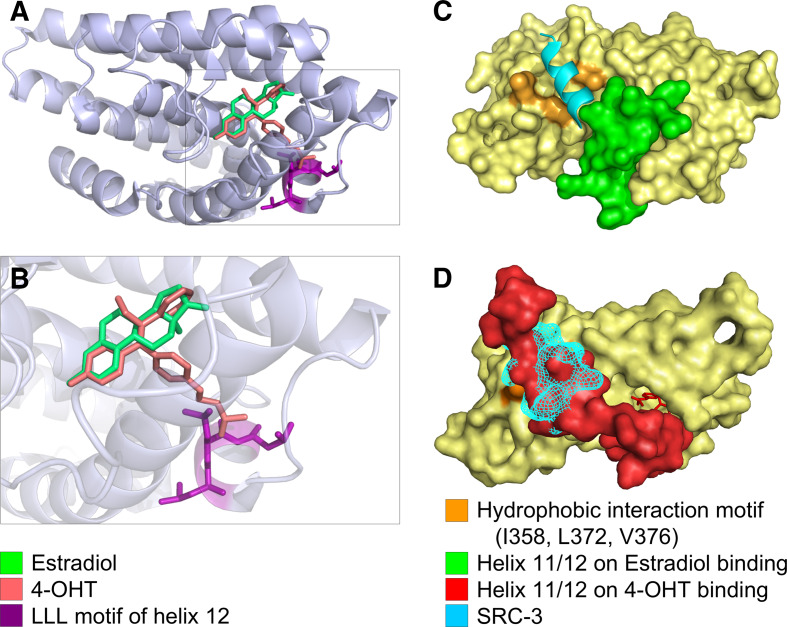

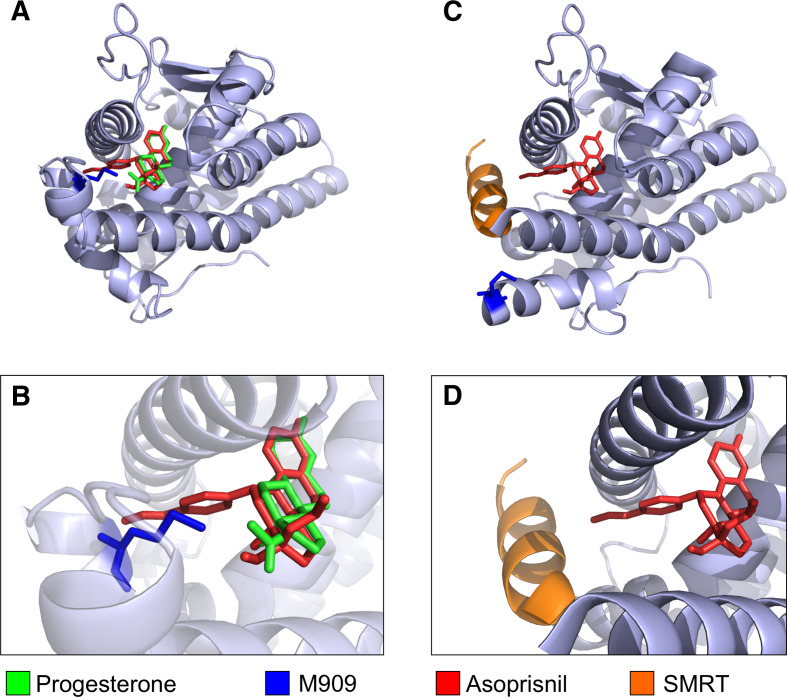
References
-
- DeLano WL. The PyMOL Molecular Graphics System. CA: Palo Alto; 2008.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
