

Identification of LRRc17 as a negative regulator of receptor activator of NF-kappaB ligand (RANKL)-induced osteoclast differentiation
- PMID: 19336404
- PMCID: PMC2685711
- DOI: 10.1074/jbc.M807722200
Identification of LRRc17 as a negative regulator of receptor activator of NF-kappaB ligand (RANKL)-induced osteoclast differentiation
Abstract
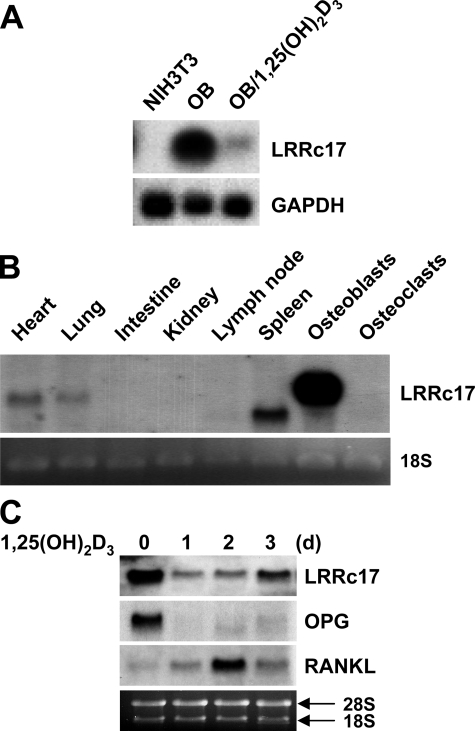
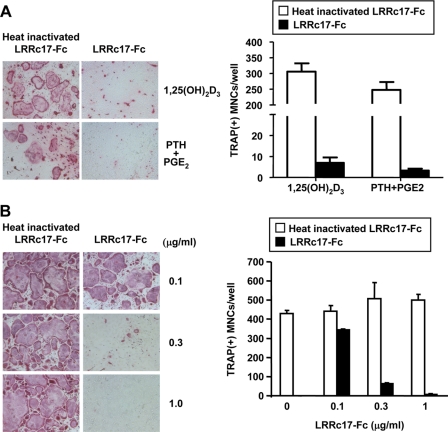
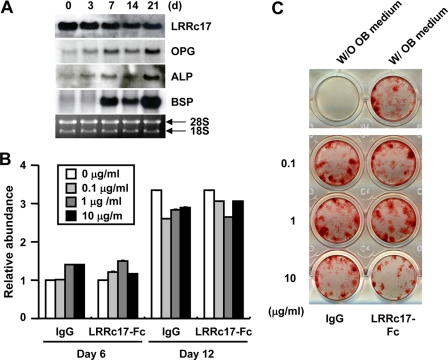
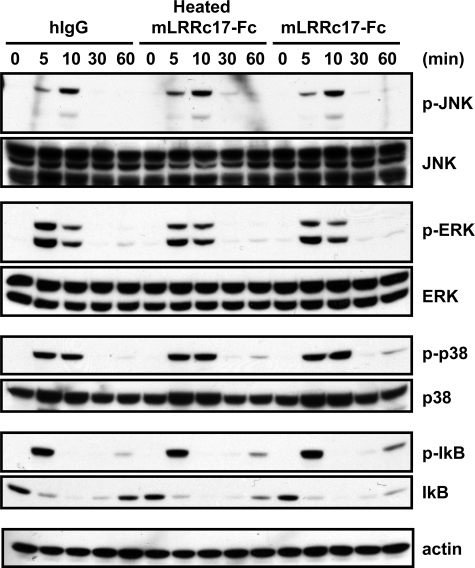
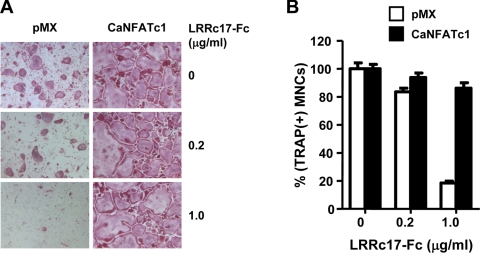
Osteoblasts are the primary cells responsible for bone formation. They also support osteoclast formation from bone marrow precursors in response to osteotropic factors by inducing receptor activator of NF-kappaB ligand (RANKL) expression and down-regulating osteoprotegerin (OPG) production. In addition to the RANKL-RANK-OPG signaling axis, other factors produced by osteoblasts/stromal cells are involved in osteoclastogenesis. Here, we describe the identification and characterization of leucine-rich repeat-containing 17 (LRRc17), a member of the LRR superfamily that acts as a negative regulator of RANKL-induced osteoclast differentiation. Osteoblasts showed high levels of LRRc17 expression, which was down-regulated in response to the pro-osteoclastogenic factor 1,25-dihydroxyvitamin D(3). Recombinant LRRc17 protein inhibited RANKL-induced osteoclast differentiation from bone marrow precursors, whereas it did not affect the differentiation or activation of macrophages and dendritic cells. These results suggest that among the cell types derived from common myeloid precursors, LRRc17 specifically regulates osteoclasts. Further analysis revealed that LRRc17 attenuated RANKL-induced expression of NFATc1 by blocking phospholipase C-gamma signaling, which, in turn, inhibited RANKL-mediated osteoclast differentiation. Taken together, our results demonstrated a novel inhibitory activity of LRRc17 in RANKL-induced osteoclastogenesis.
Figures


Similar articles
-
Coptisine inhibits RANKL-induced NF-κB phosphorylation in osteoclast precursors and suppresses function through the regulation of RANKL and OPG gene expression in osteoblastic cells.J Nat Med. 2012 Jan;66(1):8-16. doi: 10.1007/s11418-011-0537-7. Epub 2011 Jun 9. J Nat Med. 2012. PMID: 21656335
-
CTRP3 acts as a negative regulator of osteoclastogenesis through AMPK-c-Fos-NFATc1 signaling in vitro and RANKL-induced calvarial bone destruction in vivo.Bone. 2015 Oct;79:242-51. doi: 10.1016/j.bone.2015.06.011. Epub 2015 Jun 21. Bone. 2015. PMID: 26103094
-
Sophorae Flos extract inhibits RANKL-induced osteoclast differentiation by suppressing the NF-κB/NFATc1 pathway in mouse bone marrow cells.BMC Complement Altern Med. 2017 Mar 23;17(1):164. doi: 10.1186/s12906-016-1550-x. BMC Complement Altern Med. 2017. PMID: 28335757 Free PMC article.
-
Action of RANKL and OPG for osteoclastogenesis.Crit Rev Eukaryot Gene Expr. 2009;19(1):61-72. doi: 10.1615/critreveukargeneexpr.v19.i1.30. Crit Rev Eukaryot Gene Expr. 2009. PMID: 19191757 Review.
-
Osteoclast differentiation by RANKL and OPG signaling pathways.J Bone Miner Metab. 2021 Jan;39(1):19-26. doi: 10.1007/s00774-020-01162-6. Epub 2020 Oct 20. J Bone Miner Metab. 2021. PMID: 33079279 Review.
Cited by
-
Secretion of a truncated osteopetrosis-associated transmembrane protein 1 (OSTM1) mutant inhibits osteoclastogenesis through down-regulation of the B lymphocyte-induced maturation protein 1 (BLIMP1)-nuclear factor of activated T cells c1 (NFATc1) axis.J Biol Chem. 2014 Dec 26;289(52):35868-81. doi: 10.1074/jbc.M114.589614. Epub 2014 Oct 30. J Biol Chem. 2014. PMID: 25359771 Free PMC article.
-
Regulation of bone formation and remodeling by G-protein-coupled receptor 48.Development. 2009 Aug;136(16):2747-56. doi: 10.1242/dev.033571. Epub 2009 Jul 15. Development. 2009. PMID: 19605502 Free PMC article.
-
Phospholipases of mineralization competent cells and matrix vesicles: roles in physiological and pathological mineralizations.Int J Mol Sci. 2013 Mar 1;14(3):5036-129. doi: 10.3390/ijms14035036. Int J Mol Sci. 2013. PMID: 23455471 Free PMC article.
-
Advancing Biomarker Discovery and Therapeutic Targets in Duchenne Muscular Dystrophy: A Comprehensive Review.Int J Mol Sci. 2024 Jan 3;25(1):631. doi: 10.3390/ijms25010631. Int J Mol Sci. 2024. PMID: 38203802 Free PMC article. Review.
-
Construction of an artificial neural network diagnostic model and investigation of immune cell infiltration characteristics for idiopathic pulmonary fibrosis.BMC Pulm Med. 2024 Sep 17;24(1):458. doi: 10.1186/s12890-024-03249-6. BMC Pulm Med. 2024. PMID: 39289672 Free PMC article.
References
-
- Lacey, D. L., Timms, E., Tan, H. L., Kelley, M. J., Dunstan, C. R., Burgess, T., Elliott, R., Colombero, A., Elliott, G., Scully, S., Hsu, H., Sullivan, J., Hawkins, N., Davy, E., Capparelli, C., Eli, A., Qian, Y. X., Kaufman, S., Sarosi, I., Shalhoub, V., Senaldi, G., Guo, J., Delaney, J., and Boyle, W. J. (1998) Cell 93 165-176 - PubMed
-
- Suda, T., Takahashi, N., Udagawa, N., Jimi, E., Gillespie, M. T., and Martin, T. J. (1999) Endocr. Rev. 20 345-357 - PubMed
-
- Teitelbaum, S. L. (2000) Science 289 1504-1508 - PubMed
-
- Walsh, M. C., Kim, N., Kadono, Y., Rho, J., Lee, S. Y., Lorenzo, J., and Choi, Y. (2006) Annu. Rev. Immunol. 24 33-63 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
