

Inverted duplications on acentric markers: mechanism of formation
- PMID: 19336476
- PMCID: PMC2685760
- DOI: 10.1093/hmg/ddp160
Inverted duplications on acentric markers: mechanism of formation
Abstract
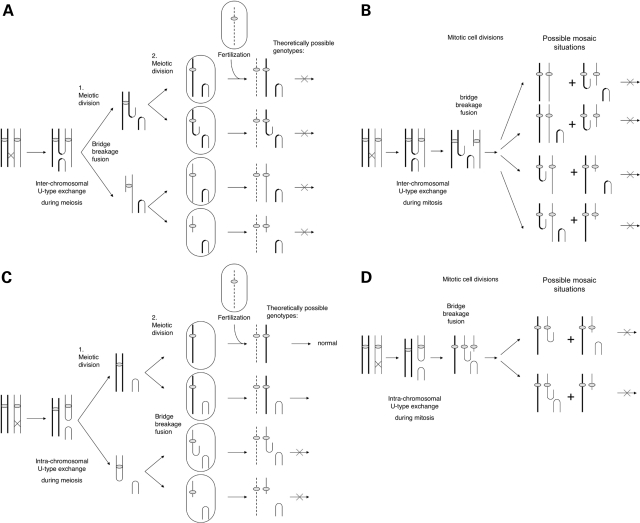
Acentric inverted duplication (inv dup) markers, the largest group of chromosomal abnormalities with neocentromere formation, are found in patients both with idiopathic mental retardation and with cancer. The mechanism of their formation has been investigated by analyzing the breakpoints and the genotypes of 12 inv dup marker cases (three trisomic, six tetrasomic, two polysomic and one X chromosome derived marker) using a combination of fluorescence in situ hybridization, quantitative SNP array and microsatellite analysis. Inv dup markers were found to form either symmetrically with one breakpoint or asymmetrically with two distinct breakpoints. Genotype analyses revealed that all inv dup markers formed from one single chromatid end. This observation is incompatible with the previously suggested model by which the acentric inv dup markers form through inter-chromosomal U-type exchange. On the basis of the identification of DNA sequence motifs with inverted homologies within all observed breakpoint regions, a new general mechanism is proposed for the acentric inv dup marker formation: following a double-strand break an acentric fragment forms, during either meiosis or mitosis. The open DNA end of the acentric fragment is stabilized by the formation of an intra-chromosomal loop promoted by the presence of sequences with inverted homologies. Likely coinciding with the neocentromere formation, this stabilized fragment is duplicated during an early mitotic event, insuring the marker's survival during cell division and its presence in all cells.
Figures






References
-
- Italiano A., Attias R., Aurias A., Perot G., Burel-Vandenbos F., Otto J., Venissac N., Pedeutour F. Molecular cytogenetic characterization of a metastatic lung sarcomatoid carcinoma: 9p23 neocentromere and 9p23–p24 amplification including JAK2 and JMJD2C. Cancer Genet. Cytogenet. 2006;167:122–130. - PubMed
-
- Abeliovich D., Yehuda O., Ben-Neriah S., Kapelushnik Y., Ben-Yehuda D. dup(10q) lacking alpha-satellite DNA in bone marrow cells of a patient with acute myeloid leukemia. Cancer Genet. Cytogenet. 1996;89:1–6. - PubMed
-
- Sirvent N., Forus A., Lescaut W., Burel F., Benzaken S., Chazal M., Bourgeon A., Vermeesch J.R., Myklebost O., Turc-Carel C., et al. Characterization of centromere alterations in liposarcomas. Genes Chrom. Cancer. 2000;29:117–129. - PubMed
-
- Gisselsson D., Hoglund M., Mertens F., Mitelman F., Mandahl N. Chromosomal organization of amplified chromosome 12 sequences in mesenchymal tumors detected by fluorescence in situ hybridization. Genes Chrom. Cancer. 1998;23:203–212. - PubMed
