

Guanylyl cyclase C prevents colon cancer metastasis by regulating tumor epithelial cell matrix metalloproteinase-9
- PMID: 19336567
- PMCID: PMC2688960
- DOI: 10.1158/0008-5472.CAN-09-0067
Guanylyl cyclase C prevents colon cancer metastasis by regulating tumor epithelial cell matrix metalloproteinase-9
Abstract
Matrix metalloproteinase-9 (MMP-9) produced by colorectal cancer cells is a critical determinant of metastatic disease progression and an attractive target for antimetastatic strategies to reduce colon cancer mortality. Cellular signaling by cyclic GMP (cGMP) regulates MMP-9 dynamics in various cell systems, and the bacterial enterotoxin receptor guanylyl cyclase C (GCC), the principle source of cGMP in colonocytes, which is overexpressed in colorectal cancers, inhibits tumor initiation and progression in the intestine. Here, we show that ligand-dependent GCC signaling through cGMP induces functional remodeling of cancer cell MMP-9 reflected by a compartmental redistribution of this gelatinase, in which intracellular retention resulted in reciprocal extracellular depletion. Functional remodeling of MMP-9 by GCC signaling reduced the ability of colon cancer cells to degrade matrix components, organize the actin cytoskeleton to form locomotory organelles and spread, and hematogenously seed distant organs. Of significance, GCC effects on cancer cell MMP-9 prevented establishment of metastatic colonies by colorectal cancer cells in the mouse peritoneum in vivo. Because endogenous hormones for GCC are uniformly deficient in intestinal tumors, reactivation of dormant GCC signaling with exogenous administration of GCC agonists may represent a specific intervention to target MMP-9 functions in colon cancer cells. The notion that GCC-mediated regulation of cancer cell MMP-9 disrupts metastasis, in turn, underscores the unexplored utility of GCC hormone replacement therapy in the chemoprevention of colorectal cancer progression.
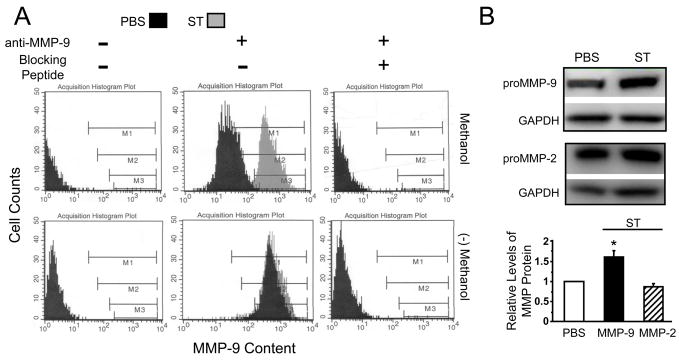
Figures





References
-
- American Cancer Society Cancer facts & figures 2008. Atlanta: American Cancer Society; 2008.
-
- Sobrero A, Kerr D, Glimelius B, et al. New directions in the treatment of colorectal cancer: a look to the future. Eur J Cancer. 2000;36:559–66. - PubMed
-
- Meyerhardt JA, Mayer RJ. Systemic therapy for colorectal cancer. N Engl J Med. 2005;352:476–87. - PubMed
-
- Jemal A, Tiwari RC, Murray T, et al. Cancer statistics, 2004. CA Cancer J Clin. 2004;54:8–29. - PubMed
-
- Fidler IJ. The pathogenesis of cancer metastasis: the ‘seed and soil’ hypothesis revisited. Nat Rev Cancer. 2003;3:453–8. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
