

What's new in neuroimaging methods?
- PMID: 19338512
- PMCID: PMC2716071
- DOI: 10.1111/j.1749-6632.2009.04420.x
What's new in neuroimaging methods?
Abstract
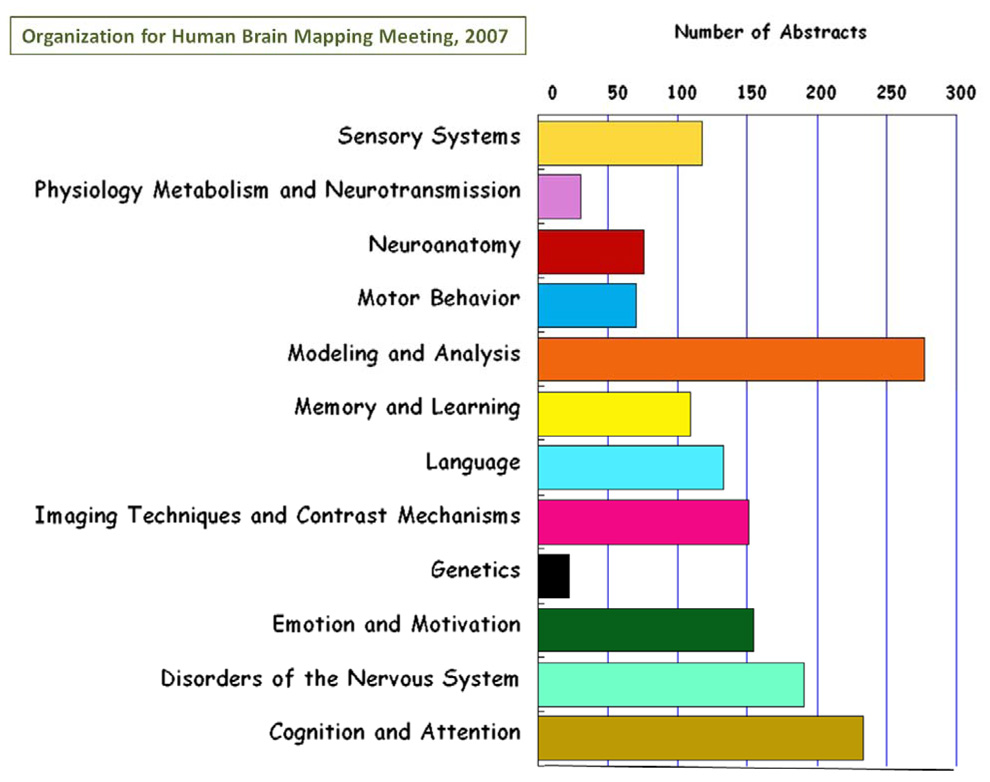
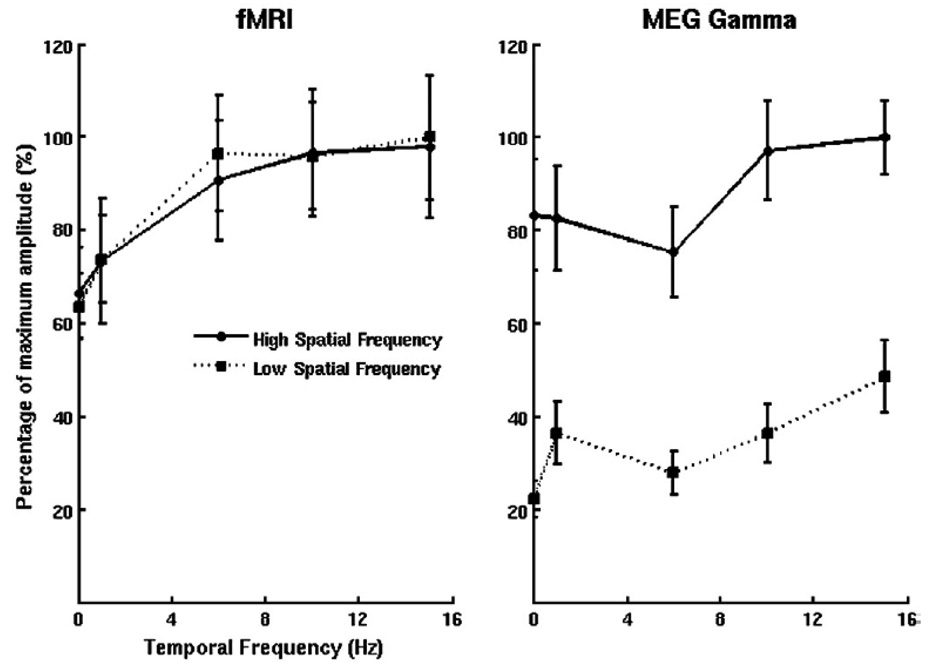
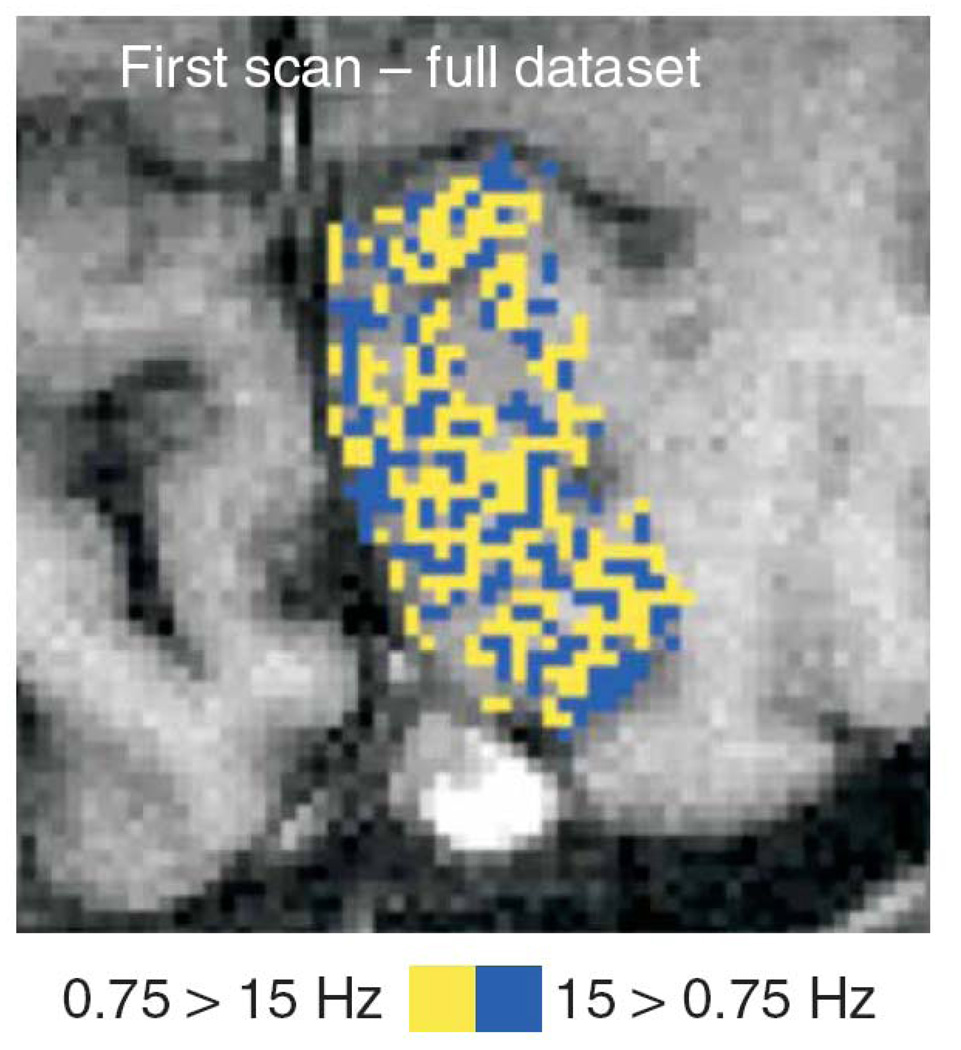
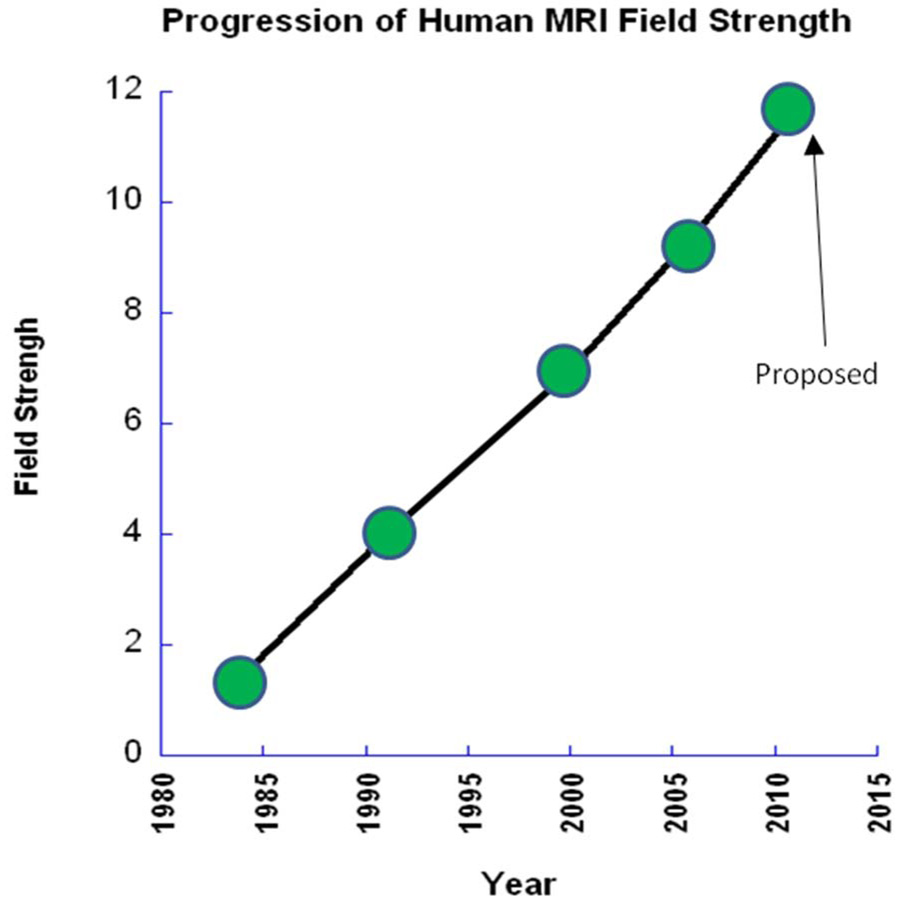
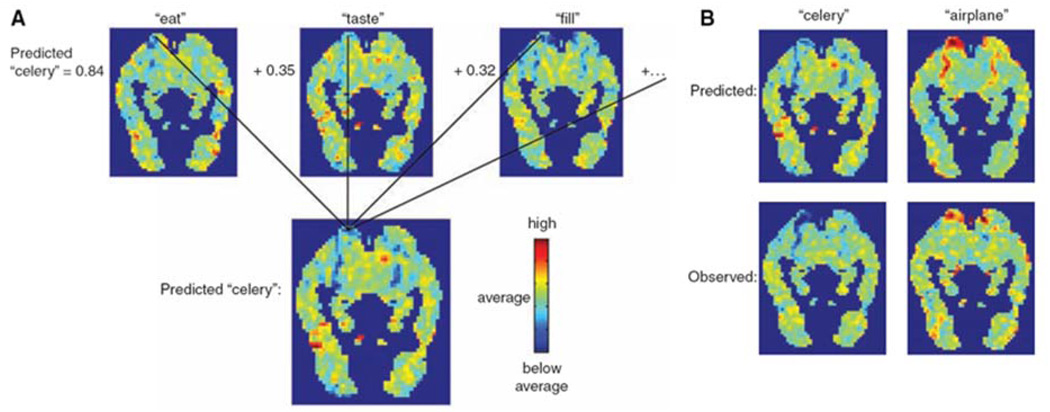
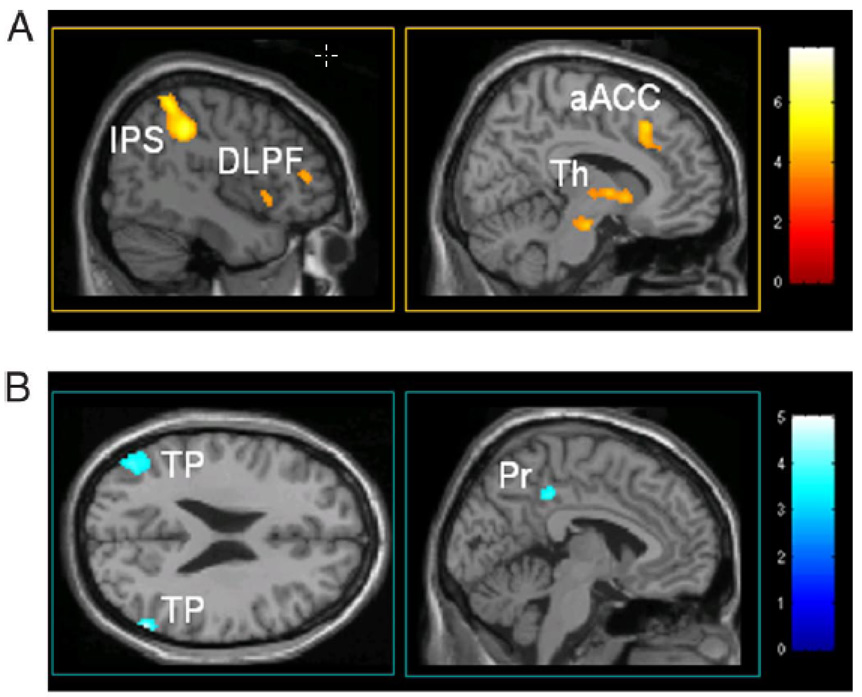
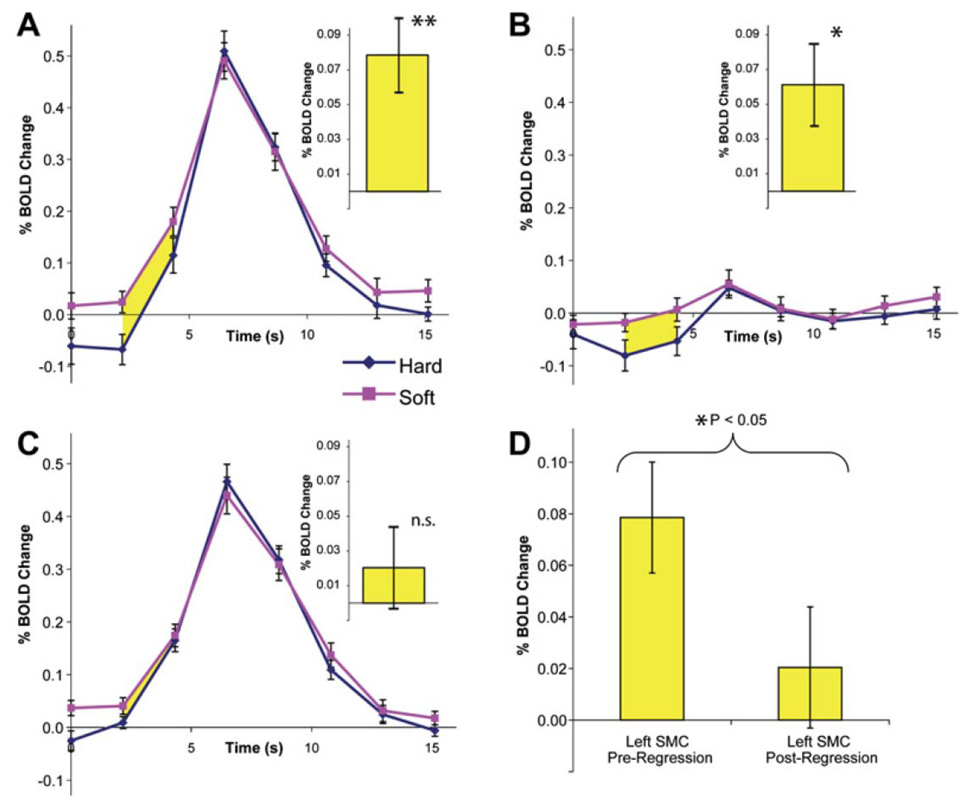
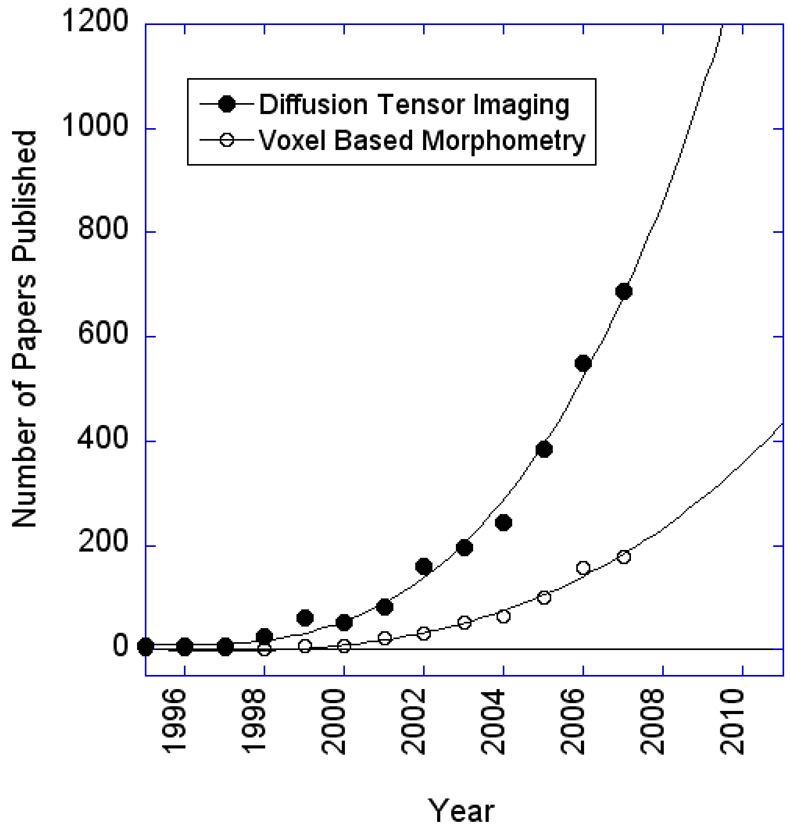
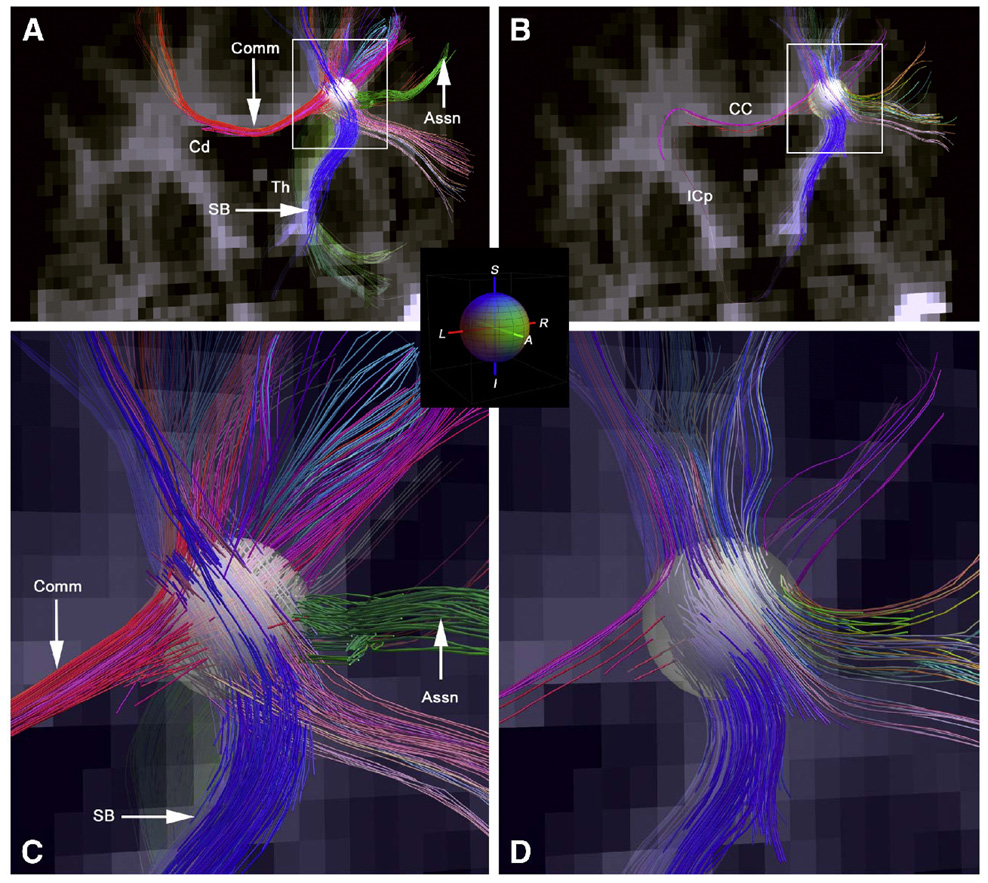
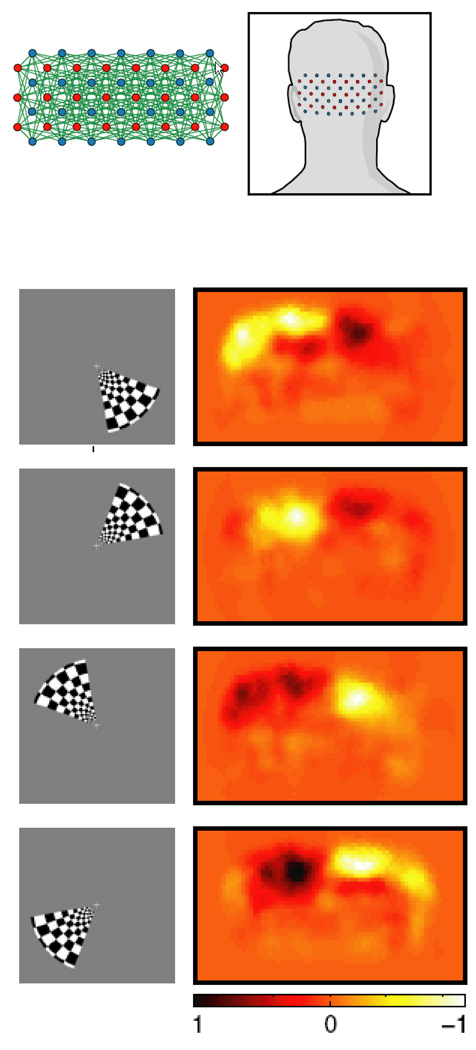
The rapid advancement of neuroimaging methodology and its growing availability has transformed neuroscience research. The answers to many questions that we ask about how the brain is organized depend on the quality of data that we are able to obtain about the locations, dynamics, fluctuations, magnitudes, and types of brain activity and structural changes. In this review an attempt is made to take a snapshot of the cutting edge of a small component of the very rapidly evolving field of neuroimaging. For each area covered, a brief context is provided along with a summary of a few of the current developments and issues. Then, several outstanding papers, published in the past year or so, are described, providing an example of the directions in which each area is progressing. The areas covered include functional magnetic resonance imaging (fMRI), voxel-based morphometry (VBM), diffusion tensor imaging (DTI), electroencephalography (EEG), magnetoencephalography (MEG), optical imaging, and positron emission tomography (PET). More detail is included on fMRI; its subsections include fMRI interpretation, new fMRI contrasts, MRI technology, MRI paradigms and processing, and endogenous oscillations in fMRI.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
