

The virological synapse facilitates herpes simplex virus entry into T cells
- PMID: 19339346
- PMCID: PMC2687377
- DOI: 10.1128/JVI.02163-08
The virological synapse facilitates herpes simplex virus entry into T cells
Abstract
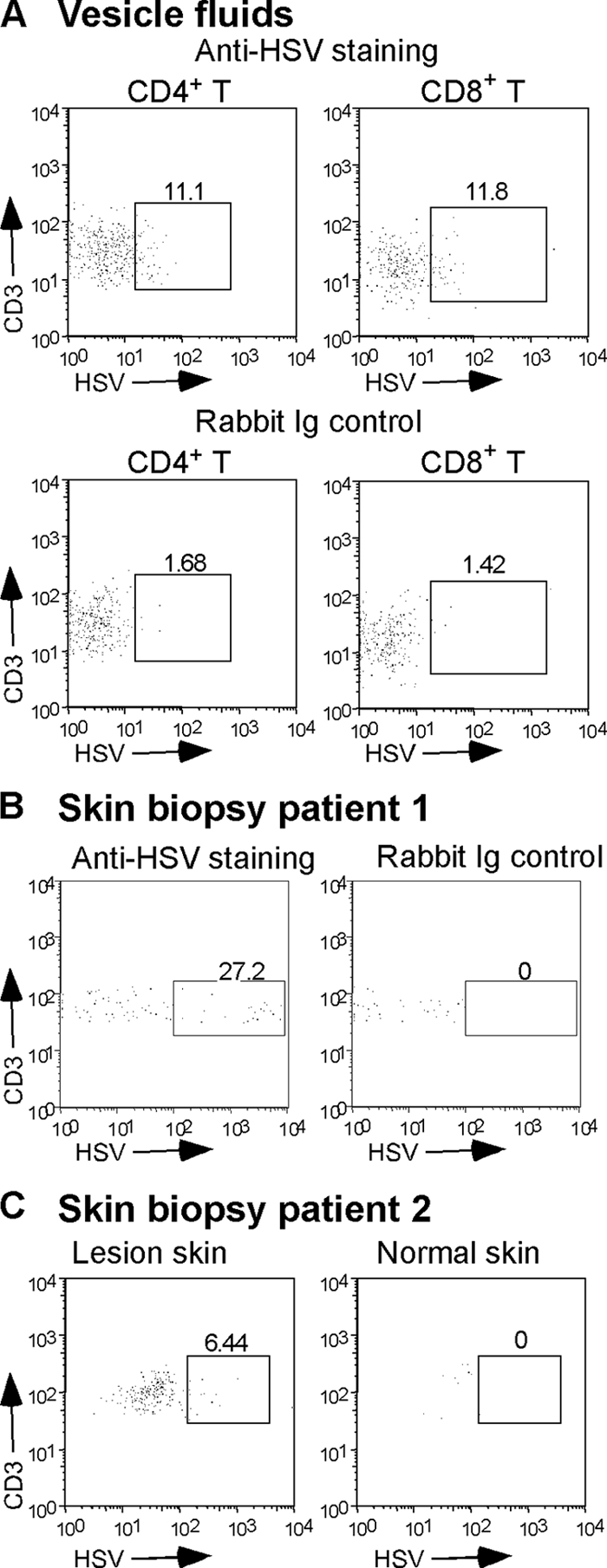
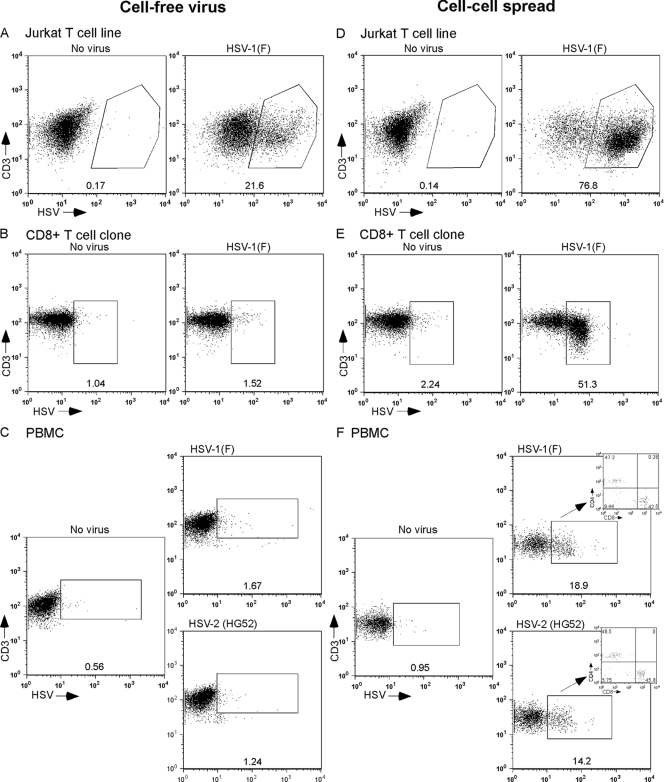
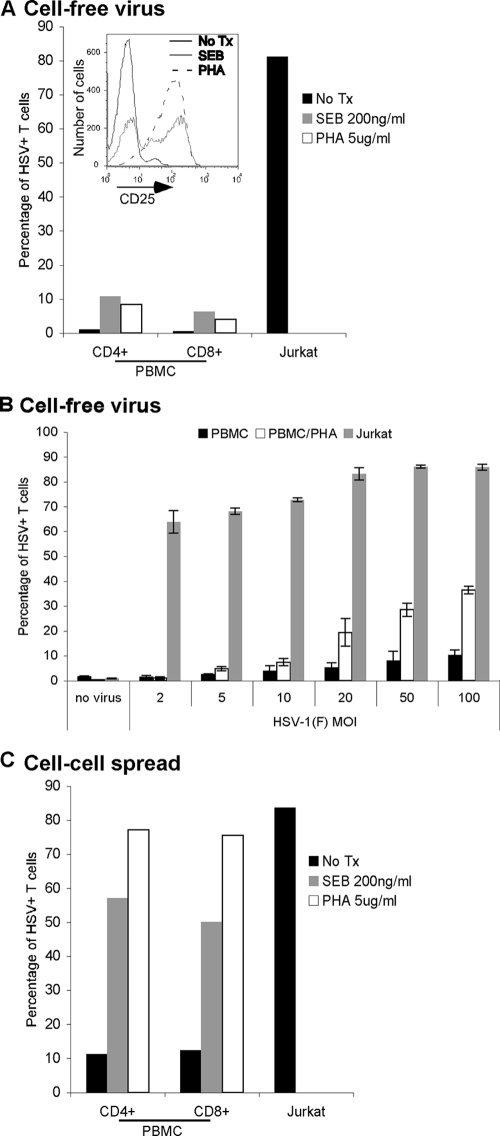
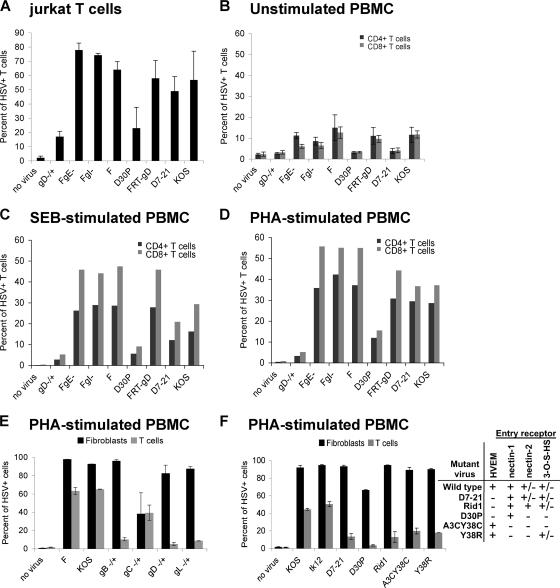
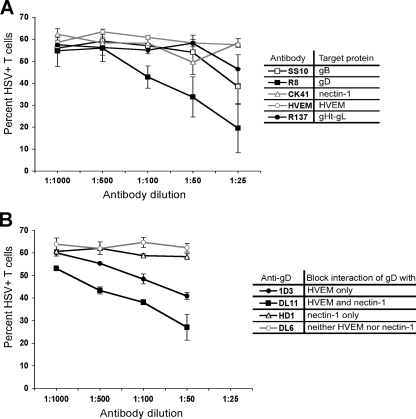
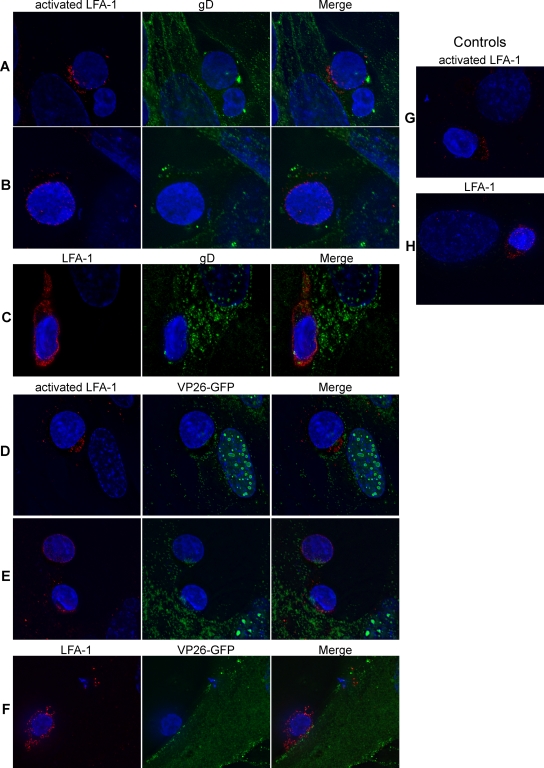
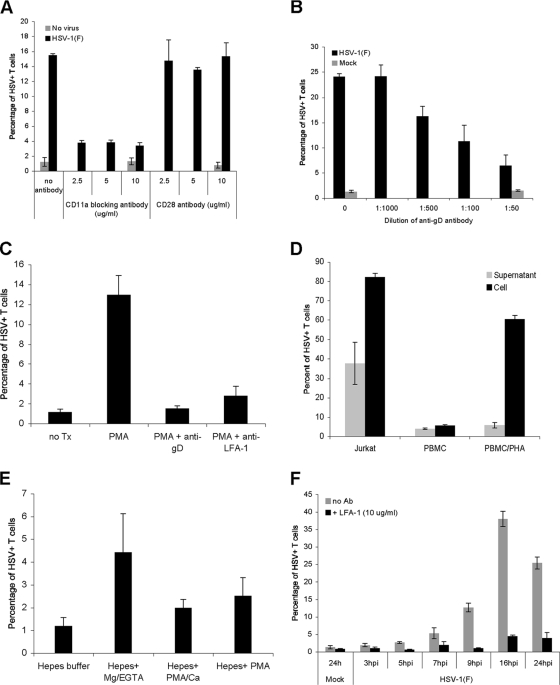
The virological synapse (VS) is a specialized molecular structure that facilitates the transfer of certain lymphotropic viruses into uninfected T cells. However, the role of the VS in the transfer of nonlymphotropic viruses into T cells is unknown. Herpes simplex virus (HSV) has been shown in vitro to infect T cells and modulate T-cell receptor function, thereby suppressing T-cell antiviral function. However, whether such infection of T cells occurs in vivo is unknown. Here, we examined whether T-cell infection could be observed in human HSV disease and investigated the mechanism of HSV entry into T cells. We found that HSV-infected T cells were readily detectable during human disease, suggesting that infection and modulation of T-cell function plays a role in human immunopathology. HSV infection of both CD4(+) and CD8(+) T cells occurred much more efficiently via direct cell-to-cell spread from infected fibroblasts than by cell-free virus. Activation of T cells increased their permissivity to HSV infection. Cell-to-cell spread to T cells did not require HSV glycoproteins E and I (gE and gI), which are critical for cell-to-cell spread between epithelial cells. Transfer of HSV to T cells required gD, and the four known entry receptors appear to be contributing to viral entry, with a dominant role for the herpesvirus entry mediator and nectin-1. VS-like structures enriched in activated lymphocyte function-associated antigen 1 (LFA-1) were observed at the point of contact between HSV-infected fibroblasts and T cells. Consistent with spread occurring via the VS, transfer of HSV was increased by activation of LFA-1, and cell-to-cell spread could be inhibited by antibodies to LFA-1 or gD. Taken together, these results constitute the first demonstration of VS-dependent cell-to-cell spread for a predominantly nonlymphotropic virus. Furthermore, they support an important role for infection and immunomodulation of T cells in clinical human disease. Targeting of the VS might allow selective immunopotentiation during infections with HSV or other nonlymphotropic viruses.
Figures
References
-
- Arthos, J., C. Cicala, E. Martinelli, K. Macleod, D. Van Ryk, D. Wei, Z. Xiao, T. D. Veenstra, T. P. Conrad, R. A. Lempicki, S. McLaughlin, M. Pascuccio, R. Gopaul, J. McNally, C. C. Cruz, N. Censoplano, E. Chung, K. N. Reitano, S. Kottilil, D. J. Goode, and A. S. Fauci. 2008. HIV-1 envelope protein binds to and signals through integrin α4β7, the gut mucosal homing receptor for peripheral T cells. Nat. Immunol. 9301-309. - PubMed
-
- Aubert, M., E. M. Krantz, and K. R. Jerome. 2006. Herpes simplex virus genes Us3, Us5, and Us12 differentially regulate cytotoxic T lymphocyte-induced cytotoxicity. Viral Immunol. 19391-408. - PubMed
-
- Bouayyad, A., and J. Menezes. 1990. Comparative study of herpes simplex virus receptor expression on human lymphoid cells. Virology 179905-910. - PubMed
-
- Brickner, A. G., E. H. Warren, J. A. Caldwell, Y. Akatsuka, T. N. Golovina, A. L. Zarling, J. Shabanowitz, L. C. Eisenlohr, D. F. Hunt, V. H. Engelhard, and S. R. Riddell. 2001. The immunogenicity of a new human minor histocompatibility antigen results from differential antigen processing. J. Exp. Med. 193195-206. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
