

Activation of antibacterial autophagy by NADPH oxidases
- PMID: 19339495
- PMCID: PMC2664152
- DOI: 10.1073/pnas.0811045106
Activation of antibacterial autophagy by NADPH oxidases
Abstract
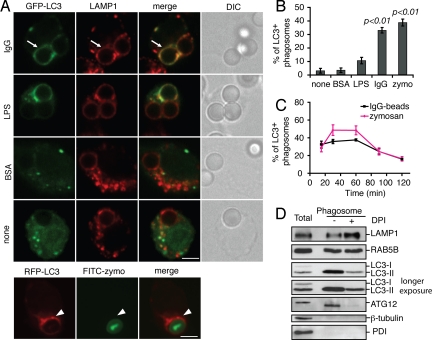
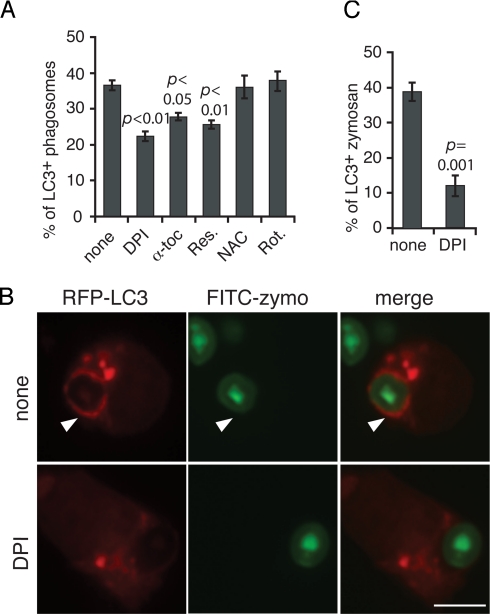
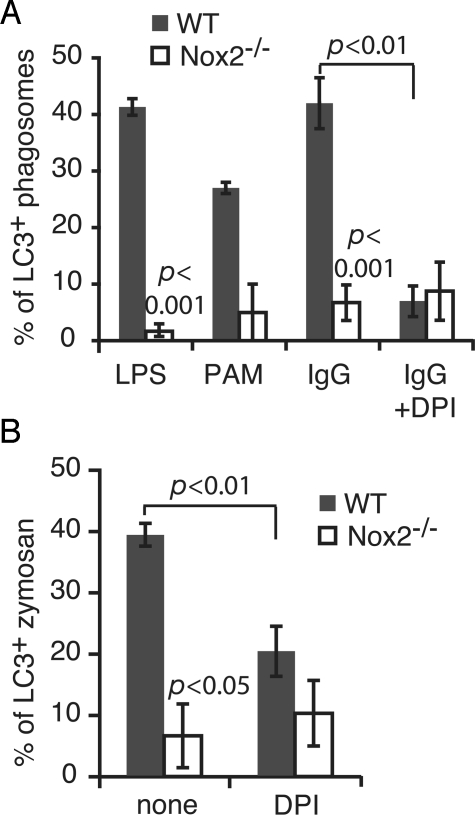
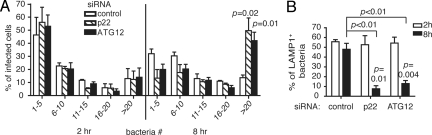
Autophagy plays an important role in immunity to microbial pathogens. The autophagy system can target bacteria in phagosomes, promoting phagosome maturation and preventing pathogen escape into the cytosol. Recently, Toll-like receptor (TLR) signaling from phagosomes was found to initiate their targeting by the autophagy system, but the mechanism by which TLR signaling activates autophagy is unclear. Here we show that autophagy targeting of phagosomes is not exclusive to those containing TLR ligands. Engagement of either TLRs or the Fcgamma receptors (FcgammaRs) during phagocytosis induced recruitment of the autophagy protein LC3 to phagosomes with similar kinetics. Both receptors are known to activate the NOX2 NADPH oxidase, which plays a central role in microbial killing by phagocytes through the generation of reactive oxygen species (ROS). We found that NOX2-generated ROS are necessary for LC3 recruitment to phagosomes. Antibacterial autophagy in human epithelial cells, which do not express NOX2, was also dependent on ROS generation. These data reveal a coupling of oxidative and nonoxidative killing activities of the NOX2 NADPH oxidase in phagocytes through autophagy. Furthermore, our results suggest a general role for members of the NOX family in regulating autophagy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Park JB. Phagocytosis induces superoxide formation and apoptosis in macrophages. Exp Mol Med. 2003;35:325–335. - PubMed
-
- Sanjuan MA, et al. Toll-like receptor signalling in macrophages links the autophagy pathway to phagocytosis. Nature. 2007;450:1253–1257. - PubMed
-
- Nakatogawa H, Ichimura Y, Ohsumi Y. Atg8, a ubiquitin-like protein required for autophagosome formation, mediates membrane tethering and hemifusion. Cell. 2007;130:165–178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
