

FXYD1 phosphorylation in vitro and in adult rat cardiac myocytes: threonine 69 is a novel substrate for protein kinase C
- PMID: 19339511
- PMCID: PMC2692419
- DOI: 10.1152/ajpcell.00523.2008
FXYD1 phosphorylation in vitro and in adult rat cardiac myocytes: threonine 69 is a novel substrate for protein kinase C
Abstract
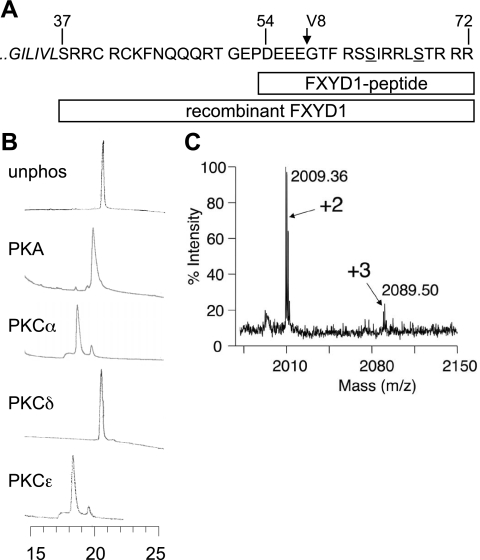
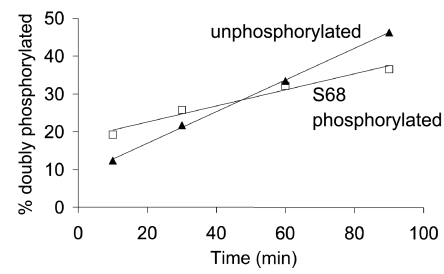
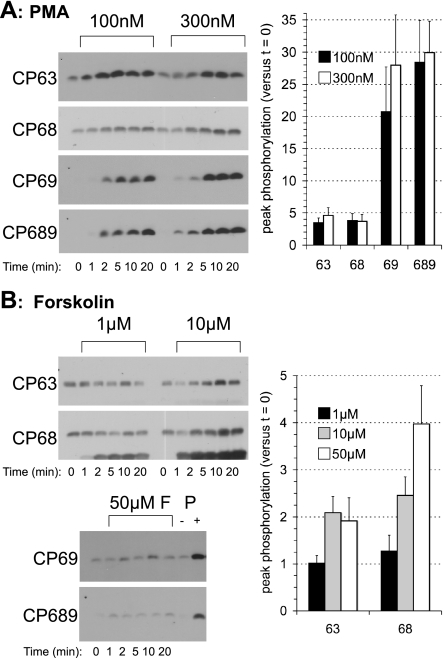
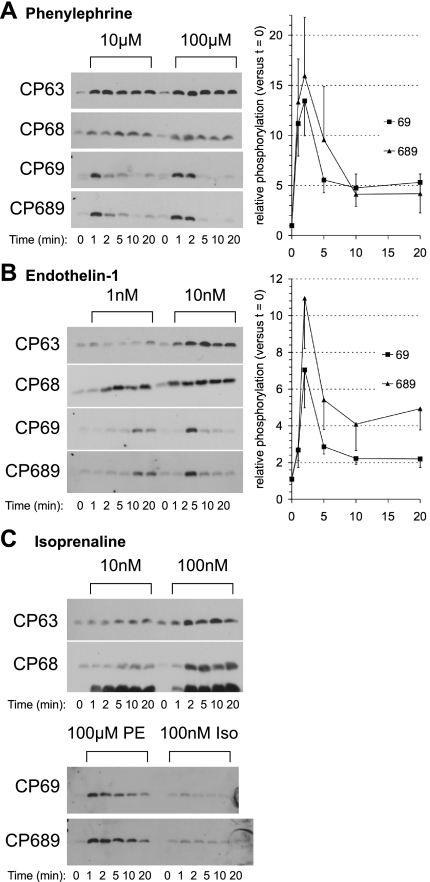
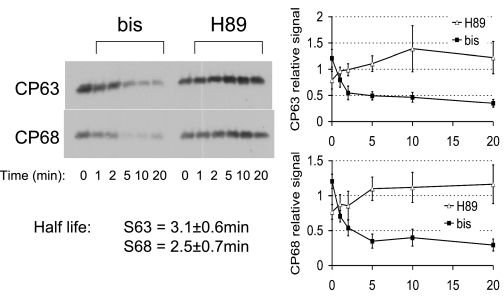
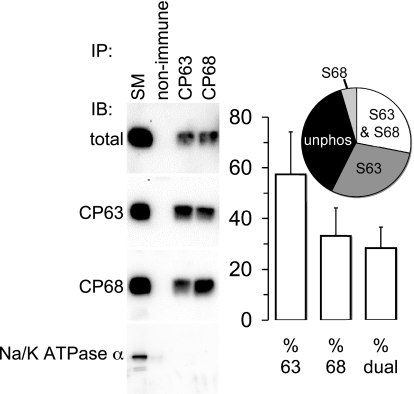
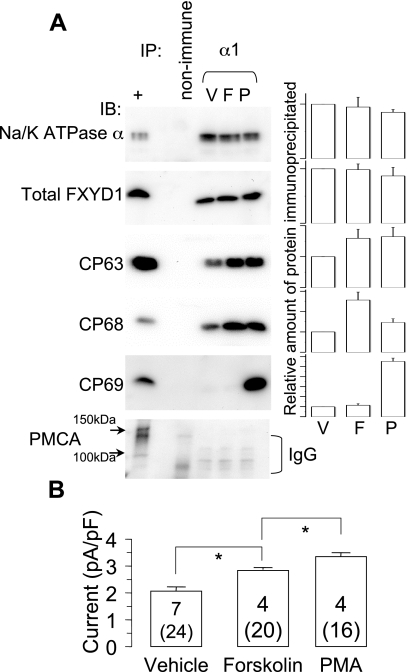
FXYD1 (phospholemman), the primary sarcolemmal kinase substrate in the heart, is a regulator of the cardiac sodium pump. We investigated phosphorylation of FXYD1 peptides by purified kinases using HPLC, mass spectrometry, and Edman sequencing, and FXYD1 phosphorylation in cultured adult rat ventricular myocytes treated with PKA and PKC agonists by phosphospecific immunoblotting. PKA phosphorylates serines 63 and 68 (S63 and S68) and PKC phosphorylates S63, S68, and a new site, threonine 69 (T69). In unstimulated myocytes, FXYD1 is approximately 30% phosphorylated at S63 and S68, but barely phosphorylated at T69. S63 and S68 are rapidly dephosphorylated following acute inhibition of PKC in unstimulated cells. Receptor-mediated PKC activation causes sustained phosphorylation of S63 and S68, but transient phosphorylation of T69. To characterize the effect of T69 phosphorylation on sodium pump function, we measured pump currents using whole cell voltage clamping of cultured adult rat ventricular myocytes with 50 mM sodium in the patch pipette. Activation of PKA or PKC increased pump currents (from 2.1 +/- 0.2 pA/pF in unstimulated cells to 2.9 +/- 0.1 pA/pF for PKA and 3.4 +/- 0.2 pA/pF for PKC). Following kinase activation, phosphorylated FXYD1 was coimmunoprecipitated with sodium pump alpha(1)-subunit. We conclude that T69 is a previously undescribed phosphorylation site in FXYD1. Acute T69 phosphorylation elicits stimulation of the sodium pump additional to that induced by S63 and S68 phosphorylation.
Figures

References
-
- Bers DM, Barry WH, Despa S. Intracellular Na+ regulation in cardiac myocytes. Cardiovasc Res 57: 897–912, 2003. - PubMed
-
- Bossuyt J, Ai X, Moorman JR, Pogwizd SM, Bers DM. Expression and phosphorylation of the Na-pump regulatory subunit phospholemman in heart failure. Circ Res 97: 558–565, 2005. - PubMed
-
- Bossuyt J, Despa S, Martin JL, Bers DM. Phospholemman phosphorylation alters its fluorescence resonance energy transfer with the Na/K-ATPase pump. J Biol Chem 281: 32765–32773, 2006. - PubMed
-
- Chibalin AV, Pedemonte CH, Katz AI, Feraille E, Berggren PO, Bertorello AM. Phosphorylation of the catalyic α-subunit constitutes a triggering signal for Na+,K+-ATPase endocytosis. J Biol Chem 273: 8814–8819, 1998. - PubMed
-
- Clerk A, Bogoyevitch MA, Anderson MB, Sugden PH. Differential activation of protein kinase C isoforms by endothelin-1 and phenylephrine and subsequent stimulation of p42 and p44 mitogen-activated protein kinases in ventricular myocytes cultured from neonatal rat hearts. J Biol Chem 269: 32848–32857, 1994. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases