

Cyclic mechanical stress and trabecular meshwork cell contractility
- PMID: 19339745
- PMCID: PMC2753281
- DOI: 10.1167/iovs.08-2694
Cyclic mechanical stress and trabecular meshwork cell contractility
Abstract
Purpose: Ocular pulse decreases outflow facility of perfused anterior segments. However, the mechanism by which conventional outflow tissues respond to cyclic intraocular pressure oscillations is unknown. The purpose of the present study was to examine responses of trabecular meshwork (TM) cells to cyclic biomechanical stress in the presence and absence of compounds known to affect cell contractility.
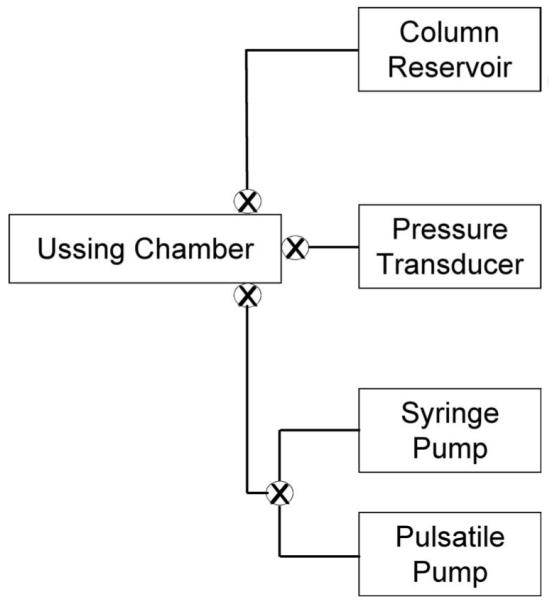
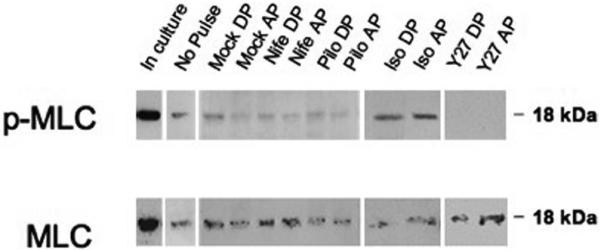
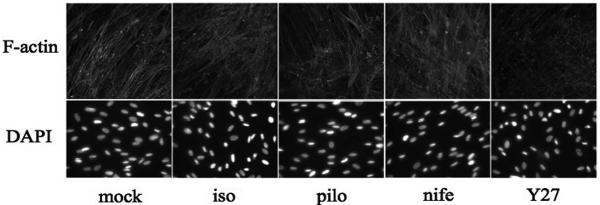
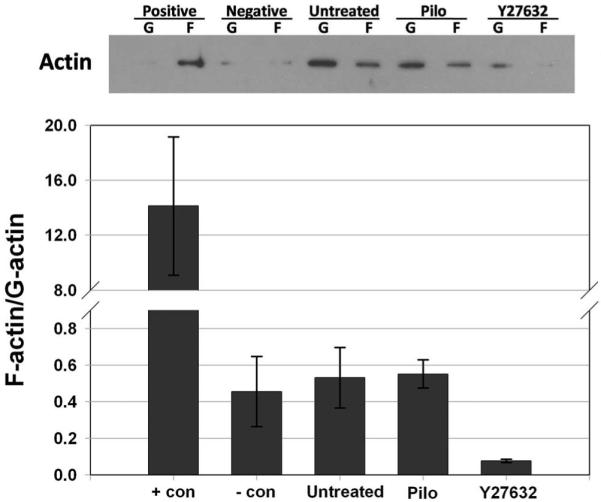
Methods: To model flow in the juxtacanalicular region of the TM and to measure changes in transendothelial flow, human TM cell monolayers on permeable filters were perfused at a constant flow rate until reaching a stable baseline pressure and then were exposed to cyclic stress with an average amplitude of 2.7 mm Hg peak to peak at a 1-Hz frequency for 2 hours in the presence or absence of compounds known to affect cell contractility (isoproterenol, Y27632, pilocarpine, and nifedipine). Pressure was recorded continuously. Immunocytochemistry staining was used to determine filamentous actin stress fiber content, whereas Western blot analysis was used to measure the extent of myosin light chain (p-MLC) phosphorylation and ratio of filamentous to globular actin.
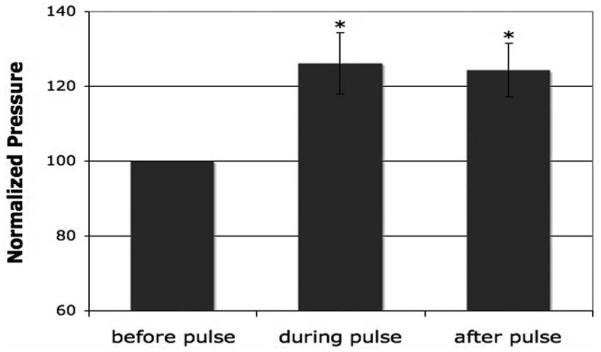
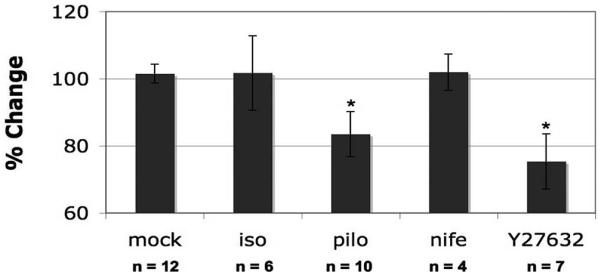
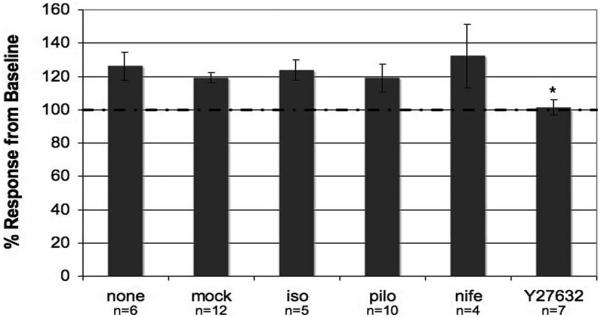
Results: Human TM cells respond to cyclic pressure oscillations by increasing mean intrachamber pressure (decreasing hydraulic conductivity) (126.13% +/- 2.4%; P < 0.05), a response blocked in the presence of Y27632, a rho-kinase inhibitor (101.35 +/- 0.59; P = 0.234), but not isoproterenol, pilocarpine, or nifedipine. Although mechanical stress appeared to have no effect, Y27632 decreased phosphorylated myosin light chain, filamentous/globular actin ratio, and stress fiber formation in TM cells.
Conclusions: Human TM cells respond to cyclic mechanical stress by increasing intrachamber pressure. Pulse-mediated effects are blocked by Y27632, implicating a role for Rho-kinase-mediated signaling and cellular contractility in ocular pulse-associated changes in outflow facility.
Figures
References
-
- Hashimoto JM, Epstein DL. Influence of intraocular pressure on aqueous outflow facility in enucleated eyes of different mammals. Invest Ophthalmol Vis Sci. 1980;19:1483–1489. - PubMed
-
- Johnstone MA. Pressure-dependent changes in nuclei and the process origins of the endothelial cells lining Schlemm’s canal. Invest Ophthalmol Vis Sci. 1979;18:44–51. - PubMed
-
- Gonzalez P, Epstein DL, Borras T. Genes upregulated in the human trabecular meshwork in response to elevated intraocular pressure. Invest Ophthalmol Vis Sci. 2000;41:352–361. - PubMed
-
- Borras T, Rowlette LL, Tamm ER, Gottanka J, Epstein DL. Effects of elevated intraocular pressure on outflow facility and TIGR/MYOC expression in perfused human anterior segments. Invest Ophthalmol Vis Sci. 2002;43:33–40. - PubMed
-
- Bradley JM, Kelley MJ, Rose A, Acott TS. Signaling pathways used in trabecular matrix metalloproteinase response to mechanical stretch. Invest Ophthalmol Vis Sci. 2003;44:5174–5181. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
