

Review
doi: 10.1038/nrm2668.
Epub 2009 Apr 2.
Ion channels versus ion pumps: the principal difference, in principle
Affiliations
- PMID: 19339978
- PMCID: PMC2742554
- DOI: 10.1038/nrm2668
Item in Clipboard
Review
Ion channels versus ion pumps: the principal difference, in principle
Nat Rev Mol Cell Biol.
2009 May.
Abstract
The incessant traffic of ions across cell membranes is controlled by two kinds of border guards: ion channels and ion pumps. Open channels let selected ions diffuse rapidly down electrical and concentration gradients, whereas ion pumps labour tirelessly to maintain the gradients by consuming energy to slowly move ions thermodynamically uphill. Because of the diametrically opposed tasks and the divergent speeds of channels and pumps, they have traditionally been viewed as completely different entities, as alike as chalk and cheese. But new structural and mechanistic information about both of these classes of molecular machines challenges this comfortable separation and forces its re-evaluation.
Figures

a/, Schematic representation of an ion channel as membrane-spanning pore through which movement of ions (red circles) is controlled by a single gate, cartooned here as a hinged door. b/, Ion pump as membrane-spanning pore with two gates that open and close alternately. Coupling of an energy source to switch the relative binding affinity for red vs. blue ions between the left-and right-hand states enables active exchange of red for blue ions across the membrane. c/, Occluded states, with both gates closed around bound ions, preclude inadvertent opening of the second gate before closure of the first gate, which would otherwise allow ions to flow down their electrochemical potential gradient several orders of magnitude faster than the pump can move them against that gradient; pumped ion movement is rate limited by the gating reactions, rather than by electrodiffusion. [Modified with permission from refs. 65, 84 ]

a/ The transmembrane regions of three subunits of a KcsA K-channel tetramer (front subunit removed for clarity) show the narrow axial selectivity filter (yellow sections of α-carbon chain), containing four closely-spaced sites for K ions (green spheres), towards the extracellular side of the membrane, and a fifth K ion in a large cavity half-way across the membrane; the single gate (activation gate) is near the cytoplasmic side. Horizontal lines mark approximate membrane boundaries. [Reproduced with permission from ref. 9]. b/ Na,K-ATPase pump complex in E2P-related conformation (with MgF42- as stable phosphate mimic; pink spheres in cytoplasmic phosphorylation, P, domain), containing two Rb ions (purple spheres) occluded within the transmembrane domain. Labels indicate the catalytic α-subunit (with grey membrane-spanning, blue phosphorylation, red nucleotide-binding, and yellow actuator, domains), auxiliary β-subunit (green), and regulatory γ-subunit (red helix). [Modified with permission from ref. 20].
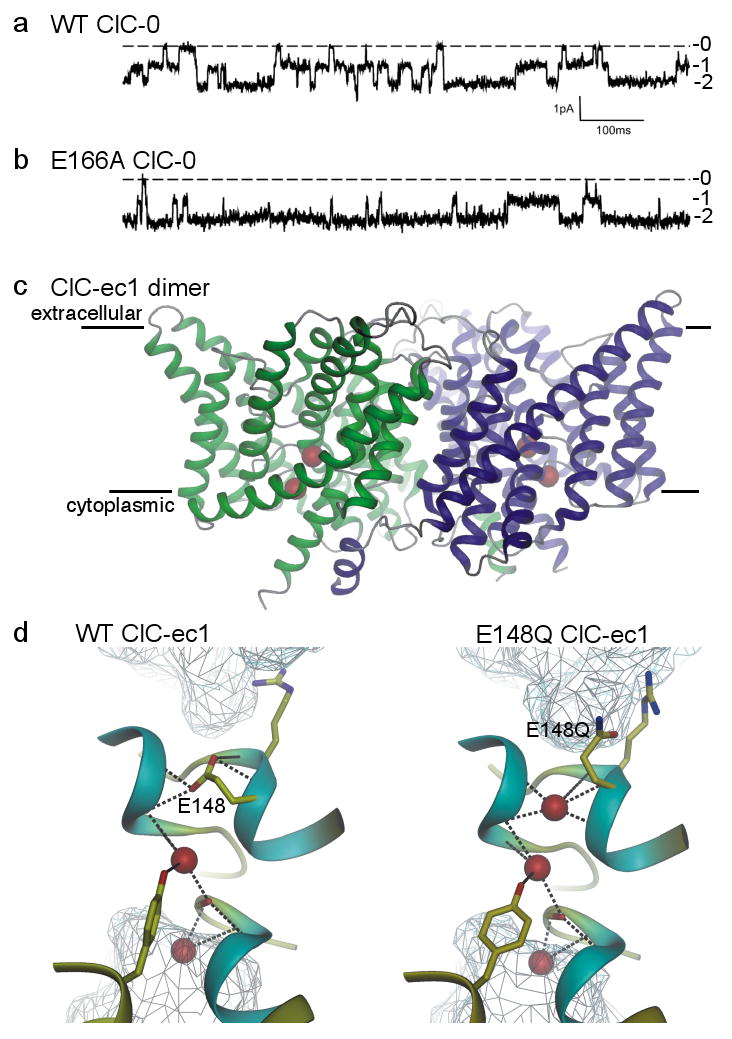
a/, b/ Current recordings showing independent opening and closing of the 2 pores in a wild-type (WT) ClC-0 channel (a), and the mostly open behaviour of an E166A ClC-0 channel (b); labels 0, 1, and 2 indicate number of open pores in each dimeric channel [Modified with permission from ref. 27]. c/ Transmembrane region of WT ClC-ec1 dimer viewed from within the membrane (membrane boundaries indicated). The ion pathway in each monomer (green, blue) contains 2 anions (red spheres). [Modified with permission from ref. 85]. d/ Pore region connecting extra- and intracellular aqueous vestibules (cyan mesh) in WT (left) and E148Q (right) ClC-ec1, containing 2 or 3 Cl ions (red spheres) respectively; the third Cl ion occupies the space vacated by the swung-out side chain of the external gate residue, 148 (E148 is equivalent to E166 of ClC-0). [Modified with permission from ref. 86].
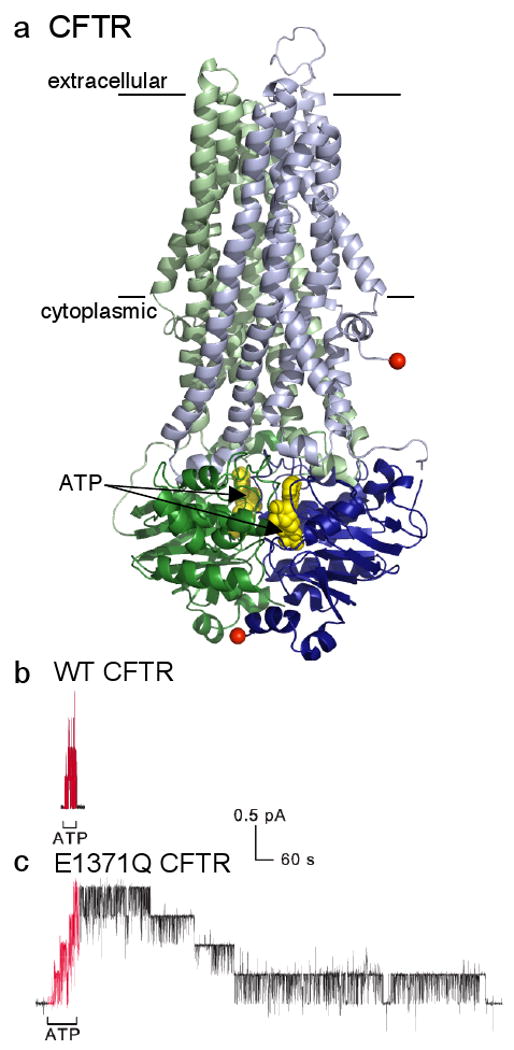
a/ Model of CFTR (N-terminal half green, C-terminal half blue) based largely on homology with prokaryotic ABC transporter, Sav1866; the transmembrane domains (lines mark membrane boundaries) and cytoplasmic linkers are lighter in colour, and the dimerized cytoplasmic nucleotide-binding domains containing 2 bound ATPs (yellow spacefill) are darker; absent is the ∼200-residue regulatory domain of unknown structure that somehow links the N- and C-terminal halves by connecting the two red spheres. [Modified with permission from ref. 87]. b/ , c/ Recordings of CFTR channel currents in excised inside-out membrane patches. Exposure to cytoplasmic ATP (red segments) allows opening and closing of WT CFTR channels (b), and of mutant E1371Q CFTR channels (c) bearing a point mutation that prevents ATP hydrolysis; each 0.4-pA current step reflects opening or closing of a single CFTR channel. The greatly delayed closing of all four E1371Q channels (c), compared to the four WT CFTR channels (b), after ATP removal shows that ATP hydrolysis times normal channel closing, and that the channel open-burst state corresponds to a conformation with ATP bound within the dimerized nucleotide-binding domains (cf. part a). [Reproduced with permission from ref. 37].
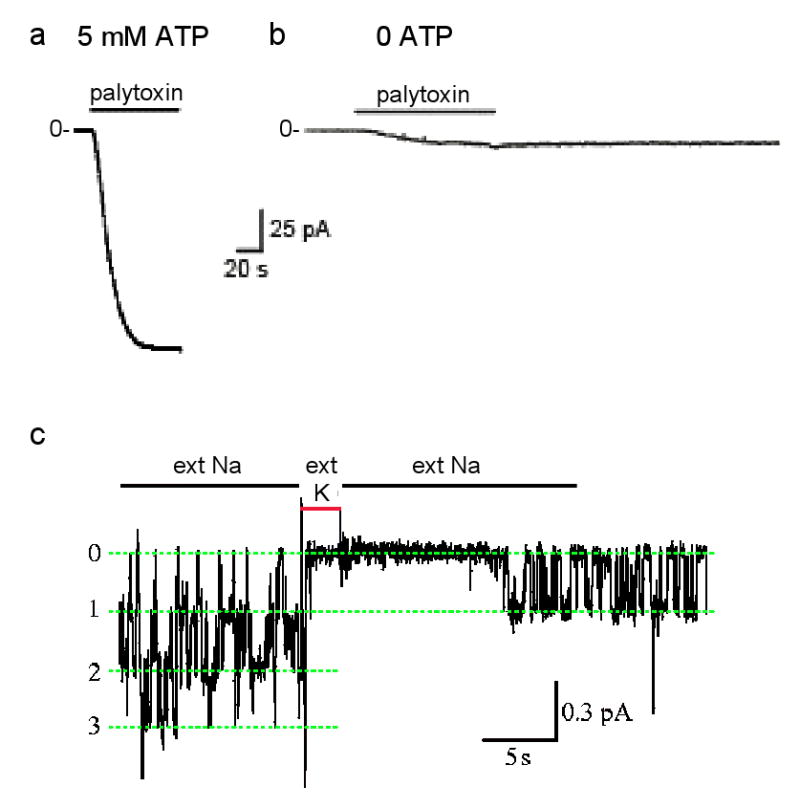
Each Na,K-ATPase pump in a cell membrane exports 3 Na ions and imports 2 K ions for each ATP hydrolyzed and can repeat this up to a hundred times a second. The net charge movement generates a current that can reach ∼20 attoamperes for a single Na,K pump, too tiny for present recording equipment. But the >20 picoamperes generated by the millions of Na,K pumps in an entire cell can be measured (e.g., ref. 79). If all K ions are removed, leaving only Na, Na,K pumping is precluded, and steady current through the pump becomes zero regardless of the membrane potential (e.g., ref. 80). This means that there is no channel-like electrodiffusive flow of Na ions through the stalled Na,K pumps, confirming that their two gates remain tightly coupled. This is the condition (zero current at -40-mV membrane potential) at the beginning of the recordings of current flow through excised outside-out patches of cell membrane each containing thousands of Na,K pumps (see figure, parts a,b). However, in presence of millimolar ATP a saturating concentration (100 nM) of palytoxin quickly elicits a relatively large current that reflects rapid Na-ion flow through Na,K pumps after palytoxin has transformed them all into cation channels,, (part a; modified with permission from ref. 81). The gates are uncoupled, but still functional Without ATP, a much weaker current is activated by the same palytoxin concentration (part b; modified with permission from ref. 81) in a comparable patch, with presumably similar numbers of palytoxin-bound Na,K pump-channels. The current is weaker because individual palytoxin-bound pump-channels spend a much greater fraction of the time closed in the absence of ATP than in its presence. This channel-opening effect of ATP is mimicked by AMPPNP or ADP, and reflects the known action of nucleotides on unmodified Na,K pumps to open the cytoplasmic-side gate,,,. External cations also modulate the probability of palytoxin-bound pump-channels being open. Just as external K ions enter unmodified Na,K pumps and become occluded in their binding sites after closure of the extracellular-side gate (Fig. 2b), brief replacement of external Na ions with K ions temporarily shuts palytoxin-bound pump-channels (part c; labels 0, 1, 2, 3 indicate number of simultaneously open pump-channels; modified with permission from ref. 65). Evidently, palytoxin transforms Na,K pumps into channels in which the two gates still respond to their physiological ligands but are no longer in synch: palytoxin breaks the fundamental rule of the one-gate-versus-two-gates formalism, which is that a pump's two gates should never be open at the same time.
References
-
- Hille B. Ion channels of excitable membranes. Vol. 814. Sinauer; Sunderland, MA: 2001.
-
- Sakmann B, Neher E. Single-Channel Recording. Plenum; New York: 1995.
-
- Läuger P. A channel mechanism for electrogenic ion pumps. Biochim Biophys Acta. 1979;552:143–161. - PubMed
-
- Patlak CS. Contributions to the theory of active transport. II. The gate-type non-carrier mechanism and generalization concerning tracer glow efficiency, and measurement of energy expenditure. Bull Math Biophys. 1957;19:209–235.
-
- Vidaver GA. Inhibition of parallel flux and augmentation of counter flux shown by transport models not involving a mobile carrier. J Theor Biol. 1966;10:301–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
